You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Vaccine Delivery TechnologyMethods and Protocols - 1071607944 PDFDocument595 pagesVaccine Delivery TechnologyMethods and Protocols - 1071607944 PDFSzeghalmi Attila100% (1)
- Anti-Stress Cem - Tech by MichaelDocument30 pagesAnti-Stress Cem - Tech by MichaelTulhina AndreiNo ratings yet
- Morrell POS review notes on sutures and inflammatory mediatorsDocument129 pagesMorrell POS review notes on sutures and inflammatory mediatorsdarrentse100% (1)
- Aqa A-Level Biology Cheatsheet PDFDocument17 pagesAqa A-Level Biology Cheatsheet PDFLiza Khan100% (6)
- Diseases and Immunity: Defence Against MicrobesDocument55 pagesDiseases and Immunity: Defence Against MicrobesIvan Prasetya100% (1)
- 3-d Immune Colored LectureDocument33 pages3-d Immune Colored LecturerohitNo ratings yet
- Zeolites Applications in Veterinary MedicineDocument17 pagesZeolites Applications in Veterinary MedicineCamelia TulcanNo ratings yet
- Immunology Serology ReviewDocument211 pagesImmunology Serology ReviewRachel Marie M. Gania100% (1)
- Akdr PPDocument1 pageAkdr PPDini AzikaNo ratings yet
- Akdr PPDocument1 pageAkdr PPDini AzikaNo ratings yet
- 19 - New Therapeutic Approach Diphenyl Diselenide Reduces Mitochondrial Dysfunction in Acetaminophen-Induced Acute Liver FailureDocument13 pages19 - New Therapeutic Approach Diphenyl Diselenide Reduces Mitochondrial Dysfunction in Acetaminophen-Induced Acute Liver FailureDini AzikaNo ratings yet
- 20 - Advances in The Management of Acute Liver FailureDocument10 pages20 - Advances in The Management of Acute Liver FailureDini AzikaNo ratings yet
- 15 - Acute Liver Failure Occurring During The First Trimester of Pregnancy Successfully Treated With Living Donor Liver TransplantationDocument4 pages15 - Acute Liver Failure Occurring During The First Trimester of Pregnancy Successfully Treated With Living Donor Liver TransplantationDini AzikaNo ratings yet
- 03 - Recent Developments in Acute Liver FailureDocument20 pages03 - Recent Developments in Acute Liver FailureDini AzikaNo ratings yet
- 15 - Acute Liver Failure Occurring During The First Trimester of Pregnancy Successfully Treated With Living Donor Liver TransplantationDocument4 pages15 - Acute Liver Failure Occurring During The First Trimester of Pregnancy Successfully Treated With Living Donor Liver TransplantationDini AzikaNo ratings yet
- Vestibuler NeuritisDocument8 pagesVestibuler NeuritisDini AzikaNo ratings yet
- 19 - New Therapeutic Approach Diphenyl Diselenide Reduces Mitochondrial Dysfunction in Acetaminophen-Induced Acute Liver FailureDocument13 pages19 - New Therapeutic Approach Diphenyl Diselenide Reduces Mitochondrial Dysfunction in Acetaminophen-Induced Acute Liver FailureDini AzikaNo ratings yet
- Print JurnalDocument0 pagesPrint JurnalDini AzikaNo ratings yet
- An Update On The Management of Acne VulgarisDocument6 pagesAn Update On The Management of Acne VulgarisDini AzikaNo ratings yet
- MouseDocument1 pageMouseDini AzikaNo ratings yet
- 3.pendrik Tandean-Interpretasi EKGDocument24 pages3.pendrik Tandean-Interpretasi EKGDini AzikaNo ratings yet
- Print JurnalDocument0 pagesPrint JurnalDini AzikaNo ratings yet
- 3.pendrik Tandean-Interpretasi EKGDocument24 pages3.pendrik Tandean-Interpretasi EKGDini AzikaNo ratings yet
- An Update On The Management of Acne VulgarisDocument6 pagesAn Update On The Management of Acne VulgarisDini AzikaNo ratings yet
- Immunological Tolerance: UnresponsivenessDocument31 pagesImmunological Tolerance: UnresponsivenessBasher BasherNo ratings yet
- T CellDocument19 pagesT CellSachin AnandNo ratings yet
- Immune Reconstitution Inflammatory Syndrome in HivDocument37 pagesImmune Reconstitution Inflammatory Syndrome in Hivpokhara gharipatanNo ratings yet
- White Blood CellDocument8 pagesWhite Blood Cellsai calderNo ratings yet
- National Dental College and Hospital: Seminar On ImmunityDocument24 pagesNational Dental College and Hospital: Seminar On Immunitydr parveen bathlaNo ratings yet
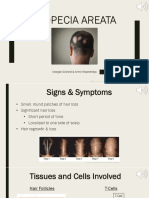
- Alopecia Areata: Morgan Gilcrest & Armir ShperdhejaDocument12 pagesAlopecia Areata: Morgan Gilcrest & Armir ShperdhejaMorgan GilcrestNo ratings yet
- Lymphatic System Anatomy and Physiology - NurseslabsDocument35 pagesLymphatic System Anatomy and Physiology - NurseslabsAlyssum MarieNo ratings yet
- 43Document15 pages43raynahollanderNo ratings yet
- PathologyDocument48 pagesPathologyAjay DivvelaNo ratings yet
- Chapter 21 - ImmunityDocument77 pagesChapter 21 - ImmunityAk62442100% (1)
- Microbes and Infection: Matthew K. Wong, Philip Barbulescu, Bryan Coburn, Elaine Reguera-Nu NezDocument9 pagesMicrobes and Infection: Matthew K. Wong, Philip Barbulescu, Bryan Coburn, Elaine Reguera-Nu NezSpore FluxNo ratings yet
- Micp Lec (Week - 4) Host Response To InfectionDocument15 pagesMicp Lec (Week - 4) Host Response To Infectioncaitie miracleNo ratings yet
- Summary Notes - Topic 6 Edexcel (A) Biology A-LevelDocument8 pagesSummary Notes - Topic 6 Edexcel (A) Biology A-LevelmohammedNo ratings yet
- Diseases of the Immune System: Hypersensitivity, Autoimmunity, ImmunodeficienciesDocument41 pagesDiseases of the Immune System: Hypersensitivity, Autoimmunity, ImmunodeficienciesFu Xiao ShanNo ratings yet
- 2 SeminarDocument39 pages2 SeminarIka Gultom100% (1)
- Lesson Plan: ObjectivesDocument4 pagesLesson Plan: ObjectivesChaden Al TawilNo ratings yet
- IL Interleukins GuideDocument10 pagesIL Interleukins GuideKhristy AbrielleNo ratings yet
- 2015 2020 Biotech Primer Quinquennial Weekly CompendiumDocument467 pages2015 2020 Biotech Primer Quinquennial Weekly CompendiumseetharamNo ratings yet
- UT Dallas Syllabus For Biol4345.001 06s Taught by John Burr (Burr)Document1 pageUT Dallas Syllabus For Biol4345.001 06s Taught by John Burr (Burr)UT Dallas Provost's Technology GroupNo ratings yet
- Gene Silencing TechniquesDocument15 pagesGene Silencing TechniquesShaher Bano Mirza100% (2)
- TCR 2+Document14 pagesTCR 2+Dome Leon TapiaNo ratings yet
- CAR T Therapy Beyond Cancer: The Evolution of A Living Drug: PerspectiveDocument9 pagesCAR T Therapy Beyond Cancer: The Evolution of A Living Drug: PerspectiveCinta DíezNo ratings yet