
A Revised Limbic System Model For Memory, Emotion and Behaviour PDF
A Revised Limbic System Model For Memory, Emotion and Behaviour PDF
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Aleksandr Romanovich Luria Auth. Higher Cortical Functions in Man PDFDocument524 pagesAleksandr Romanovich Luria Auth. Higher Cortical Functions in Man PDFSharonParedes100% (1)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Brain Bee NotesDocument10 pagesBrain Bee Notesapi-534972164No ratings yet
- Brodman MapDocument18 pagesBrodman MapBecky Edwards0% (1)
- Pub - Spanish Cooking Traditional Dishes and Regional SP PDFDocument166 pagesPub - Spanish Cooking Traditional Dishes and Regional SP PDFjason1989No ratings yet
- Music Psychology in EducationDocument289 pagesMusic Psychology in Educationworm123_12389% (9)
- The Neural Basis of Lexicon and Grammar in First and Second Language: The Declarative/procedural ModelDocument18 pagesThe Neural Basis of Lexicon and Grammar in First and Second Language: The Declarative/procedural Models0453737No ratings yet
- Math Standards PDFDocument93 pagesMath Standards PDFjason1989No ratings yet
- Recycling Household CFLs. 09 07Document5 pagesRecycling Household CFLs. 09 07jason1989No ratings yet
- Bio 1Document1 pageBio 1jason1989No ratings yet
- Heat TransferDocument37 pagesHeat Transferjason1989No ratings yet
- Volker Hoenig Stanford 2008 UploadDocument28 pagesVolker Hoenig Stanford 2008 Uploadjason1989No ratings yet
- Oily WasteDocument1 pageOily Wastejason1989No ratings yet
- Auditory Pattern Perception in 'Split Brain' PatientsDocument3 pagesAuditory Pattern Perception in 'Split Brain' PatientsAnthonyNo ratings yet
- Bab 2-08211141014Document38 pagesBab 2-08211141014Bunga frzNo ratings yet
- Ungerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFDocument18 pagesUngerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFJhamil Andre Venero LovatonNo ratings yet
- 5 MASP Neuropsychology of ReadingDocument30 pages5 MASP Neuropsychology of ReadingAnonymous 6IqMo06No ratings yet
- Neurology 1.03 Approach To Cerebral Function - Dr. MartinezDocument7 pagesNeurology 1.03 Approach To Cerebral Function - Dr. MartinezDeomicah SolanoNo ratings yet
- CVA JroozDocument5 pagesCVA JroozEdmonLagartoNo ratings yet
- Lc200 Stsrting SystemDocument6 pagesLc200 Stsrting SystemMichael Frank Vasques RenaciaNo ratings yet
- Neuro ErgonomicsDocument271 pagesNeuro ErgonomicsDedy IsmailNo ratings yet
- Unit 3 CALP Module For LENSDocument33 pagesUnit 3 CALP Module For LENSLauren Michelle RanjoNo ratings yet
- Cognitive Psychology Theory Process and Methodology 1st Edition Mcbride Test BankDocument25 pagesCognitive Psychology Theory Process and Methodology 1st Edition Mcbride Test BankAndrewMartinezmren100% (50)
- RISEN INDEED Versi IndonesiaDocument3 pagesRISEN INDEED Versi IndonesiaYuldin JimNo ratings yet
- Tbi IntroDocument50 pagesTbi IntroAjjugal SushmajaNo ratings yet
- Neurotheology - Brain, Science, Spirituality, Religious ExperienceDocument31 pagesNeurotheology - Brain, Science, Spirituality, Religious ExperienceElianKitha100% (3)
- The Brain - TRANSESDocument4 pagesThe Brain - TRANSESJamailah EncenzoNo ratings yet
- Semiologi Epilepsi: Nama Pasien: No CM: Tanggal: Diagnosis Awal: PemeriksaDocument7 pagesSemiologi Epilepsi: Nama Pasien: No CM: Tanggal: Diagnosis Awal: Pemeriksaanton MDNo ratings yet
- Temporal Lobe Epilepsy: Beate Diehl and John S. DuncanDocument4 pagesTemporal Lobe Epilepsy: Beate Diehl and John S. DuncanViviana Torres BallesterosNo ratings yet
- Assessment and Therapeutic Application of The Expressive Therapies Continuum Implications For Brain Structures and FunctionsDocument11 pagesAssessment and Therapeutic Application of The Expressive Therapies Continuum Implications For Brain Structures and FunctionsCarla Horta100% (1)
- Brain ThesisDocument6 pagesBrain ThesisHolly Fisher100% (2)
- Location - Routing Engine Compartment Wire and Wire (RHD 1GD-FTV, 2GD-FTV)Document2 pagesLocation - Routing Engine Compartment Wire and Wire (RHD 1GD-FTV, 2GD-FTV)Anibal Fabian GonzalezNo ratings yet
- The Andromedan EmpireDocument12 pagesThe Andromedan EmpireJeremyGarnerNo ratings yet
- Lobes of The BrainDocument5 pagesLobes of The BrainEnforcement SectionNo ratings yet
- Sts Cheat Sheet of The BrainDocument30 pagesSts Cheat Sheet of The BrainRahula RakeshNo ratings yet
- Fontes 2016Document9 pagesFontes 2016ivavukojaNo ratings yet
- Biological Psychology Assignment 1Document9 pagesBiological Psychology Assignment 1Saulhe ImranNo ratings yet
- The Cognitive Nueorsicence of Human Memory Since HMDocument32 pagesThe Cognitive Nueorsicence of Human Memory Since HMOscar WongNo ratings yet


































































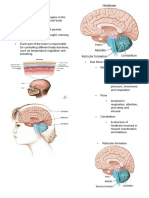







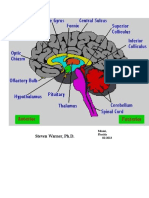



You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Aleksandr Romanovich Luria Auth. Higher Cortical Functions in Man PDFDocument524 pagesAleksandr Romanovich Luria Auth. Higher Cortical Functions in Man PDFSharonParedes100% (1)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Brain Bee NotesDocument10 pagesBrain Bee Notesapi-534972164No ratings yet
- Brodman MapDocument18 pagesBrodman MapBecky Edwards0% (1)
- Pub - Spanish Cooking Traditional Dishes and Regional SP PDFDocument166 pagesPub - Spanish Cooking Traditional Dishes and Regional SP PDFjason1989No ratings yet
- Music Psychology in EducationDocument289 pagesMusic Psychology in Educationworm123_12389% (9)
- The Neural Basis of Lexicon and Grammar in First and Second Language: The Declarative/procedural ModelDocument18 pagesThe Neural Basis of Lexicon and Grammar in First and Second Language: The Declarative/procedural Models0453737No ratings yet
- Math Standards PDFDocument93 pagesMath Standards PDFjason1989No ratings yet
- Recycling Household CFLs. 09 07Document5 pagesRecycling Household CFLs. 09 07jason1989No ratings yet
- Bio 1Document1 pageBio 1jason1989No ratings yet
- Heat TransferDocument37 pagesHeat Transferjason1989No ratings yet
- Volker Hoenig Stanford 2008 UploadDocument28 pagesVolker Hoenig Stanford 2008 Uploadjason1989No ratings yet
- Oily WasteDocument1 pageOily Wastejason1989No ratings yet
- Auditory Pattern Perception in 'Split Brain' PatientsDocument3 pagesAuditory Pattern Perception in 'Split Brain' PatientsAnthonyNo ratings yet
- Bab 2-08211141014Document38 pagesBab 2-08211141014Bunga frzNo ratings yet
- Ungerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFDocument18 pagesUngerleider & Bell (2011) - Uncovering The Visual Alphabet. Advances in Our Understanding PDFJhamil Andre Venero LovatonNo ratings yet
- 5 MASP Neuropsychology of ReadingDocument30 pages5 MASP Neuropsychology of ReadingAnonymous 6IqMo06No ratings yet
- Neurology 1.03 Approach To Cerebral Function - Dr. MartinezDocument7 pagesNeurology 1.03 Approach To Cerebral Function - Dr. MartinezDeomicah SolanoNo ratings yet
- CVA JroozDocument5 pagesCVA JroozEdmonLagartoNo ratings yet
- Lc200 Stsrting SystemDocument6 pagesLc200 Stsrting SystemMichael Frank Vasques RenaciaNo ratings yet
- Neuro ErgonomicsDocument271 pagesNeuro ErgonomicsDedy IsmailNo ratings yet
- Unit 3 CALP Module For LENSDocument33 pagesUnit 3 CALP Module For LENSLauren Michelle RanjoNo ratings yet
- Cognitive Psychology Theory Process and Methodology 1st Edition Mcbride Test BankDocument25 pagesCognitive Psychology Theory Process and Methodology 1st Edition Mcbride Test BankAndrewMartinezmren100% (50)
- RISEN INDEED Versi IndonesiaDocument3 pagesRISEN INDEED Versi IndonesiaYuldin JimNo ratings yet
- Tbi IntroDocument50 pagesTbi IntroAjjugal SushmajaNo ratings yet
- Neurotheology - Brain, Science, Spirituality, Religious ExperienceDocument31 pagesNeurotheology - Brain, Science, Spirituality, Religious ExperienceElianKitha100% (3)
- The Brain - TRANSESDocument4 pagesThe Brain - TRANSESJamailah EncenzoNo ratings yet
- Semiologi Epilepsi: Nama Pasien: No CM: Tanggal: Diagnosis Awal: PemeriksaDocument7 pagesSemiologi Epilepsi: Nama Pasien: No CM: Tanggal: Diagnosis Awal: Pemeriksaanton MDNo ratings yet
- Temporal Lobe Epilepsy: Beate Diehl and John S. DuncanDocument4 pagesTemporal Lobe Epilepsy: Beate Diehl and John S. DuncanViviana Torres BallesterosNo ratings yet
- Assessment and Therapeutic Application of The Expressive Therapies Continuum Implications For Brain Structures and FunctionsDocument11 pagesAssessment and Therapeutic Application of The Expressive Therapies Continuum Implications For Brain Structures and FunctionsCarla Horta100% (1)
- Brain ThesisDocument6 pagesBrain ThesisHolly Fisher100% (2)
- Location - Routing Engine Compartment Wire and Wire (RHD 1GD-FTV, 2GD-FTV)Document2 pagesLocation - Routing Engine Compartment Wire and Wire (RHD 1GD-FTV, 2GD-FTV)Anibal Fabian GonzalezNo ratings yet
- The Andromedan EmpireDocument12 pagesThe Andromedan EmpireJeremyGarnerNo ratings yet
- Lobes of The BrainDocument5 pagesLobes of The BrainEnforcement SectionNo ratings yet
- Sts Cheat Sheet of The BrainDocument30 pagesSts Cheat Sheet of The BrainRahula RakeshNo ratings yet
- Fontes 2016Document9 pagesFontes 2016ivavukojaNo ratings yet
- Biological Psychology Assignment 1Document9 pagesBiological Psychology Assignment 1Saulhe ImranNo ratings yet
- The Cognitive Nueorsicence of Human Memory Since HMDocument32 pagesThe Cognitive Nueorsicence of Human Memory Since HMOscar WongNo ratings yet