You might also like
- Toxins 11 00383 v2Document25 pagesToxins 11 00383 v2ungureanucameliaNo ratings yet
- insecticidal_potential_of_streptomyces_against_dengue_mosquito_Document14 pagesinsecticidal_potential_of_streptomyces_against_dengue_mosquito_AidNo ratings yet
- MetarhiziummajusDocument8 pagesMetarhiziummajusHadi AgusNo ratings yet
- Hexane Antimicrob L.pyriformeDocument5 pagesHexane Antimicrob L.pyriformealNo ratings yet
- Microbiología: Selection of Strains Toxic To Cotton Boll Weevil (Anthonomus Coleoptera: Curculionidae) LarvaeDocument9 pagesMicrobiología: Selection of Strains Toxic To Cotton Boll Weevil (Anthonomus Coleoptera: Curculionidae) Larvaewilman Martinez herreraNo ratings yet
- Jurnal Lotus 4Document7 pagesJurnal Lotus 4SITI RAHMAWATINo ratings yet
- JPAM Vol 15 Issue1 P 232-239Document8 pagesJPAM Vol 15 Issue1 P 232-239Jefri Nur HidayatNo ratings yet
- An Assignment On Insect Resistan T PlantsDocument9 pagesAn Assignment On Insect Resistan T Plantsvj88No ratings yet
- In-Vitro Inhibitory Indices of Selected Fungal Isolates Against Mycotoxin FungiDocument8 pagesIn-Vitro Inhibitory Indices of Selected Fungal Isolates Against Mycotoxin FungiOpenaccess Research paperNo ratings yet
- tmp4615 TMPDocument22 pagestmp4615 TMPFrontiersNo ratings yet
- InsectBiochemistry and Molecular Biology - 2013Document9 pagesInsectBiochemistry and Molecular Biology - 2013matiasNo ratings yet
- Genetic Enhancement of Chickpea For Pod Borer Resistance Through Expression of Cryiac ProteinDocument4 pagesGenetic Enhancement of Chickpea For Pod Borer Resistance Through Expression of Cryiac ProteinDayakar PhotosNo ratings yet
- Evaluation of The Production of Antifungal Metabolites Against MutagenesisDocument9 pagesEvaluation of The Production of Antifungal Metabolites Against MutagenesisDũng NguyễnNo ratings yet
- Biopesticidal Activity of Baccilus Thuringiensis and Its ApplicationDocument15 pagesBiopesticidal Activity of Baccilus Thuringiensis and Its Applicationfaisal97No ratings yet
- Sphaerica and Oscillatoria AgardhiiDocument17 pagesSphaerica and Oscillatoria AgardhiiMhemeydha Luphe YudhaNo ratings yet
- Wahyudi Et AlDocument7 pagesWahyudi Et AlShilajit BaruaNo ratings yet
- MCB 311 & MCB 309 New VersionDocument69 pagesMCB 311 & MCB 309 New VersionChaos HadesNo ratings yet
- Biocontrol Activity of Bacillus Amyloliquefaciens CNU114001 Against Fungal Plant DiseasesDocument10 pagesBiocontrol Activity of Bacillus Amyloliquefaciens CNU114001 Against Fungal Plant DiseasesASAADNo ratings yet
- Trichoderma Fungi ControlDocument7 pagesTrichoderma Fungi ControlJorge TurpoNo ratings yet
- Transgenic Plants For Insect ResistanceDocument11 pagesTransgenic Plants For Insect ResistanceVida Faith GalvezNo ratings yet
- Dang 2009Document13 pagesDang 2009EEDIEB Prof. Millton Marques CurvoNo ratings yet
- M.P.prasad and Sunayana DagarDocument11 pagesM.P.prasad and Sunayana DagarAbhishek SinghNo ratings yet
- Diversity and Antimicrobial Potential in Sea Anemone and Holothurian MicrobiomesDocument21 pagesDiversity and Antimicrobial Potential in Sea Anemone and Holothurian MicrobiomesAde Rizky FajrullohNo ratings yet
- ICBS Conference Proceedings Secondary MetabolitesDocument9 pagesICBS Conference Proceedings Secondary MetabolitesPhương NguyễnNo ratings yet
- Collection of Rice Tungro-Infected Plants From Hotspots in The PhilippinesDocument12 pagesCollection of Rice Tungro-Infected Plants From Hotspots in The PhilippinesGilfredo V. Legada III.No ratings yet
- Git Microbiome The Effect of Finger Millet (Eleusinecoracana) Seed Extract On Albino Wistar RatsDocument8 pagesGit Microbiome The Effect of Finger Millet (Eleusinecoracana) Seed Extract On Albino Wistar RatsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Secondary Metabolites Production by ActinomycetesDocument10 pagesSecondary Metabolites Production by ActinomycetesRujean Romy Porteza GuisandoNo ratings yet
- GMOs and Trials PDFDocument138 pagesGMOs and Trials PDFSamiha AshreenNo ratings yet
- Zenniel Et. Al., 2002Document13 pagesZenniel Et. Al., 2002Aswawi AwiNo ratings yet
- Biotechnology and Its Applications NotesDocument45 pagesBiotechnology and Its Applications NotesNarasimha Pavan BNo ratings yet
- Flores Vallejo2020Document12 pagesFlores Vallejo2020maczweiNo ratings yet
- Pujiastuti 2018 IOP Conf. Ser. Earth Environ. Sci. 102 012064Document9 pagesPujiastuti 2018 IOP Conf. Ser. Earth Environ. Sci. 102 012064indra maulanaNo ratings yet
- Published Paper 11-7 - 6025 - PDFDocument11 pagesPublished Paper 11-7 - 6025 - PDFAhmed IsmailNo ratings yet
- 1 s2.0 S0944501317306602 MainDocument18 pages1 s2.0 S0944501317306602 Mainpremanand1990No ratings yet
- Bacterial Endophytes: Recent Developments and ApplicationsDocument9 pagesBacterial Endophytes: Recent Developments and ApplicationsmaczweiNo ratings yet
- Short Communication: Antifungal Potential of Crude Extracts of Trichoderma SPPDocument5 pagesShort Communication: Antifungal Potential of Crude Extracts of Trichoderma SPPKaren Avalos VelaNo ratings yet
- 14340-Article Text-1084052-1-10-20230609Document8 pages14340-Article Text-1084052-1-10-20230609AnggunNo ratings yet
- B. Thuringiensis 4Document14 pagesB. Thuringiensis 4Laura Constanza GomezNo ratings yet
- Ehp0108 000919Document12 pagesEhp0108 000919Tothazan SandaNo ratings yet
- fmicb-10-02774Document15 pagesfmicb-10-02774Marta EstradaNo ratings yet
- Chapter 34Document22 pagesChapter 34Konishko DeyNo ratings yet
- HRMS BacillaeneDocument27 pagesHRMS BacillaeneRosyida Ismi (Rosyi)No ratings yet
- Diaporthe Endophytica and D Terebinthifolii From Medicinal - 2016 - MicrobiologDocument8 pagesDiaporthe Endophytica and D Terebinthifolii From Medicinal - 2016 - MicrobiologMaria BatoolNo ratings yet
- Biotech ResearchDocument7 pagesBiotech Researchthekingraja2602No ratings yet
- JMB 33 4 543Document9 pagesJMB 33 4 543Robert StryjakNo ratings yet
- Bacillus VelezensisDocument3 pagesBacillus VelezensismstiqueoNo ratings yet
- Engineering Pathogen Resistance in Crop Plants Current Trends and Future ProspectsDocument11 pagesEngineering Pathogen Resistance in Crop Plants Current Trends and Future ProspectsUdayendu BarikNo ratings yet
- Sorghum AntiviralDocument5 pagesSorghum Antiviralnugraha widyatmonoNo ratings yet
- USP General Chapter (1115) Bioburdin Control For Non Sterile Drug SubstanceDocument8 pagesUSP General Chapter (1115) Bioburdin Control For Non Sterile Drug SubstanceDr usama El Shafey100% (7)
- Diospiros PDFDocument6 pagesDiospiros PDFvCPNo ratings yet
- Biocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaDocument9 pagesBiocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaMaria BatoolNo ratings yet
- 2012-Bti Cry Toxin ActivityDocument10 pages2012-Bti Cry Toxin ActivityDwiNo ratings yet
- Antimicrobial Activities of Polyhexamethylene GuanidineDocument6 pagesAntimicrobial Activities of Polyhexamethylene GuanidineFranco CapeloNo ratings yet
- Tugas KompreDocument6 pagesTugas KompreNadilla islamiNo ratings yet
- Genetically Engineered Plants and Crops: Definitions: Genetic ModificationDocument3 pagesGenetically Engineered Plants and Crops: Definitions: Genetic ModificationNighWolfNo ratings yet
- B. Thuringiensis Crocidolomia Binotalis: Toxicity of Isolates From Indonesia and Philippines Against ZellDocument7 pagesB. Thuringiensis Crocidolomia Binotalis: Toxicity of Isolates From Indonesia and Philippines Against ZellHilmi Fadhil AgustianNo ratings yet
- Streptomyces - Penicillium DigitatumDocument15 pagesStreptomyces - Penicillium DigitatumOpenaccess Research paperNo ratings yet
- Characterization of New Bacterial Biocontrol Agents Acinetobacter, Grapevine Resistance Against Botrytis CinereaDocument12 pagesCharacterization of New Bacterial Biocontrol Agents Acinetobacter, Grapevine Resistance Against Botrytis CinerearicardoNo ratings yet
- Detection of Major Facilitator Superfamily (MFS) Transporter in Enterobacteriaceae Isolated From Chicken (Rizal Et Al., 2022)Document6 pagesDetection of Major Facilitator Superfamily (MFS) Transporter in Enterobacteriaceae Isolated From Chicken (Rizal Et Al., 2022)Syaiful RizalNo ratings yet
- Microbiological ResearchDocument7 pagesMicrobiological Researchwilman Martinez herreraNo ratings yet
- Characterization of Bacillus Thuringiensis Isolates by Their Insecticidal Activity and Their Production of Cry and Vip3 ProteinsDocument18 pagesCharacterization of Bacillus Thuringiensis Isolates by Their Insecticidal Activity and Their Production of Cry and Vip3 Proteinswilman Martinez herreraNo ratings yet
- Kuehniella (Zeller) and Plodia Interpunctella (Hübner) LarvaeDocument9 pagesKuehniella (Zeller) and Plodia Interpunctella (Hübner) Larvaewilman Martinez herreraNo ratings yet
- Journal Pone 0206813 PDFDocument18 pagesJournal Pone 0206813 PDFwilman Martinez herreraNo ratings yet
- Biological Control: Article InfoDocument7 pagesBiological Control: Article Infowilman Martinez herreraNo ratings yet
- Kuehniella (Zeller) and Plodia Interpunctella (Hübner) LarvaeDocument9 pagesKuehniella (Zeller) and Plodia Interpunctella (Hübner) Larvaewilman Martinez herreraNo ratings yet
- Solución Semana #2Document1 pageSolución Semana #2wilman Martinez herreraNo ratings yet
- Solución Primera Semana #1Document1 pageSolución Primera Semana #1wilman Martinez herreraNo ratings yet
- University of Waterloo Thesis RepositoryDocument5 pagesUniversity of Waterloo Thesis Repositoryafknkzkkb100% (2)
- SAP & ERP Introduction: Centralized ApplicationsDocument6 pagesSAP & ERP Introduction: Centralized ApplicationsMesumNo ratings yet
- DBQ Christianity and Latin America-1Document6 pagesDBQ Christianity and Latin America-1Duggu RagguNo ratings yet
- Role of Incentive in Building MoraleDocument2 pagesRole of Incentive in Building MoraleAmina Sadaf Ansari100% (1)
- Introduction To Logics and Critical Thinking SlidesDocument14 pagesIntroduction To Logics and Critical Thinking SlidesMKNo ratings yet
- NP 4Document4 pagesNP 4Hamad RayhanNo ratings yet
- Case Study Walking The WalkDocument3 pagesCase Study Walking The Walk9899 Mariam Akter Marketing100% (1)
- Understanding Balance of PaymentsDocument8 pagesUnderstanding Balance of PaymentsMurtaza JamaliNo ratings yet
- Demerit System PointsDocument87 pagesDemerit System PointsBusinessTech25% (4)
- Black & White 2 Prima Official Game GuideDocument160 pagesBlack & White 2 Prima Official Game GuideJoep KulderijNo ratings yet
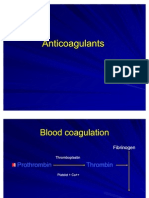
- AnticoagulantsDocument19 pagesAnticoagulantsOsama ZbedaNo ratings yet
- Sorting Lesson PlanDocument4 pagesSorting Lesson PlanStasha DuttNo ratings yet
- The Lucky Symbol of A Fish On The PalmDocument4 pagesThe Lucky Symbol of A Fish On The PalmANTHONY WRITER80% (5)
- Regents Prep 4Document7 pagesRegents Prep 4api-24284785450% (2)
- Gods Revelation in Jesus and in His ChurchDocument13 pagesGods Revelation in Jesus and in His ChurchMerry DaceraNo ratings yet
- Candombe para JoseDocument1 pageCandombe para JosestatuhominisNo ratings yet
- Harry Potter Essay 1 OutlineDocument10 pagesHarry Potter Essay 1 OutlinevickaduzerNo ratings yet
- Diogenes The Cynic and Shakespeare's Bitter FoolDocument31 pagesDiogenes The Cynic and Shakespeare's Bitter FoolGEORGE FELIPE BERNARDES BARBOSA BORGESNo ratings yet
- Acid Base ImbalanceDocument50 pagesAcid Base ImbalanceDian Pratiwi BurnamaNo ratings yet
- Séquence 4 NEW PPDDFFDocument6 pagesSéquence 4 NEW PPDDFFMohamed GuersNo ratings yet
- Marco de Luca FrontDocument1 pageMarco de Luca FrontДмитро ДуганецьNo ratings yet
- SCA No. 8220-R - Petitioner's MemorandumDocument9 pagesSCA No. 8220-R - Petitioner's MemorandumElizerNo ratings yet
- Philippines Supreme Court rules on robbery and double homicide caseDocument13 pagesPhilippines Supreme Court rules on robbery and double homicide caseNadzlah BandilaNo ratings yet
- Merchant Banking Roles and FunctionsDocument18 pagesMerchant Banking Roles and Functionssunita prabhakarNo ratings yet
- Case Nizar SummaryDocument3 pagesCase Nizar SummaryShahera Zainudin100% (1)
- Elecius Franklin IndictmentDocument2 pagesElecius Franklin Indictmentthe kingfishNo ratings yet
- CRQ Sample Questions and AnswersDocument10 pagesCRQ Sample Questions and AnswersBadda SarNo ratings yet
- Professor Weissman's Algebra Classroom 01whole NumbersDocument12 pagesProfessor Weissman's Algebra Classroom 01whole NumbersTheMathProf100% (4)
- Publishers Classification Schemes For Test ScoresDocument23 pagesPublishers Classification Schemes For Test ScoreslobotNo ratings yet
- A Case Study On SchistosomiasisDocument154 pagesA Case Study On SchistosomiasisCarmellaDawn100% (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageFrom EverandFearfully and Wonderfully: The Marvel of Bearing God's ImageRating: 5 out of 5 stars5/5 (40)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)