You might also like
- Beyond Defense: Regulation of Neuronal Morphogenesis and Brain Functions Via Toll-Like ReceptorsDocument13 pagesBeyond Defense: Regulation of Neuronal Morphogenesis and Brain Functions Via Toll-Like ReceptorsNicole MeloNo ratings yet
- The Role of Pattern-Recognition Receptors in Innate Immunity: Update On Toll-Like ReceptorsDocument12 pagesThe Role of Pattern-Recognition Receptors in Innate Immunity: Update On Toll-Like ReceptorslilianapradaNo ratings yet
- Pathogen Recognition and Innate ImmunityDocument19 pagesPathogen Recognition and Innate ImmunityMuhammad AthamnaNo ratings yet
- Pi Is 0092867410000231Document16 pagesPi Is 0092867410000231MaximeNo ratings yet
- Pathogen Recognition With Toll-Like Receptors: Taro Kawai and Shizuo AkiraDocument7 pagesPathogen Recognition With Toll-Like Receptors: Taro Kawai and Shizuo Akiraetik ainun rohmahNo ratings yet
- Pattern Recognition Receptors (PRRS) : Rossana Zaru, University of Dundee, UkDocument4 pagesPattern Recognition Receptors (PRRS) : Rossana Zaru, University of Dundee, UkFree Escort ServiceNo ratings yet
- Toll-Like Receptors and Innate ImmunityDocument14 pagesToll-Like Receptors and Innate ImmunityJUAREZ GONZALEZ EDWARD HASSANNo ratings yet
- Fimmu 05 00386Document13 pagesFimmu 05 00386Dani PazNo ratings yet
- Tollip: A Multitasking Protein in Innate Immunity and Protein TraffickingDocument8 pagesTollip: A Multitasking Protein in Innate Immunity and Protein TraffickingFadhil AhsanNo ratings yet
- The TLR Signaling Adaptor TRAM Interacts With TRAF6 To Mediate Activation of The Inflammatory Response by TLR4Document21 pagesThe TLR Signaling Adaptor TRAM Interacts With TRAF6 To Mediate Activation of The Inflammatory Response by TLR4Nadia NasrNo ratings yet
- Fimmu 13 812774Document22 pagesFimmu 13 812774nutritionist1234567No ratings yet
- Toll-Like Receptors: The Swiss Army Knife of Immunity and Vaccine DevelopmentDocument10 pagesToll-Like Receptors: The Swiss Army Knife of Immunity and Vaccine DevelopmentWilson Barros LuizNo ratings yet
- Nat Rev Immunol 2011 WestDocument15 pagesNat Rev Immunol 2011 WestGabriel CamarenaNo ratings yet
- Imm12143 (Articulo 2)Document15 pagesImm12143 (Articulo 2)LiZz LaraNo ratings yet
- Yang 2017Document10 pagesYang 2017wendyNo ratings yet
- Review Article The Role of Innate Immunity in Pulmonary InfectionsDocument14 pagesReview Article The Role of Innate Immunity in Pulmonary InfectionsAbalzNo ratings yet
- C-Type Lectin Receptor-Induced NF-KB Activation in InnateDocument8 pagesC-Type Lectin Receptor-Induced NF-KB Activation in InnateStefania CristinaNo ratings yet
- Cell InflammasomeDocument12 pagesCell InflammasomemgaborNo ratings yet
- Pattern Recognition ReceptorsDocument15 pagesPattern Recognition Receptorsratnesh gautamNo ratings yet
- Pathologic Basis of Veterinary Disease, 4th Edition: Chapter 5 Diseases of ImmunityDocument88 pagesPathologic Basis of Veterinary Disease, 4th Edition: Chapter 5 Diseases of ImmunityEdzel Tisado SacloloNo ratings yet
- TLR Wikki Part 1Document3 pagesTLR Wikki Part 1jebabalajiiiNo ratings yet
- Toll-Like Receptors (TLRS) and Nod-Like Receptors (NLRS) in InflammatoryDocument12 pagesToll-Like Receptors (TLRS) and Nod-Like Receptors (NLRS) in InflammatoryLarissa SilvaNo ratings yet
- LPS-TLR4 Tranduction PathwaysDocument7 pagesLPS-TLR4 Tranduction PathwaysGERMAN MATIAS TAPIA CURIMILNo ratings yet
- Toll-Like Receptors and Skin: JeadvDocument10 pagesToll-Like Receptors and Skin: JeadvBenor Amri MustaqimNo ratings yet
- Asst - Prof. Meroj A. Jasem Ph.D. Student Molecular Immunology CourseDocument26 pagesAsst - Prof. Meroj A. Jasem Ph.D. Student Molecular Immunology CourseahmadNo ratings yet
- Macrophage Response Towards LPSDocument10 pagesMacrophage Response Towards LPSajaewoongNo ratings yet
- Keestra Et Al 2010 tlr21Document9 pagesKeestra Et Al 2010 tlr21misilpiyapiyoNo ratings yet
- Role of Tlr2, Tlr4, and Myd88 in Murine Ozone-Induced Airway Hyperresponsiveness and NeutrophiliaDocument7 pagesRole of Tlr2, Tlr4, and Myd88 in Murine Ozone-Induced Airway Hyperresponsiveness and NeutrophiliaArkhan HanafiNo ratings yet
- Signalling of TLRsDocument8 pagesSignalling of TLRsArhi WidhiaNo ratings yet
- Sha Et Al-2004Document7 pagesSha Et Al-2004Dani PazNo ratings yet
- Learning Objectives: DR Finney The Innate Immune System, CMMB527 W22Document8 pagesLearning Objectives: DR Finney The Innate Immune System, CMMB527 W22rtm haiderNo ratings yet
- TLR Wikki 3Document3 pagesTLR Wikki 3jebabalajiiiNo ratings yet
- Avian VirusesDocument10 pagesAvian VirusesMohsan UllahNo ratings yet
- TLR DrosophilaDocument8 pagesTLR DrosophilaSaraTorresNo ratings yet
- Bacterial 23S LibreDocument2 pagesBacterial 23S LibreMuhammadRifa'iAl-FarabiNo ratings yet
- Toll-Like Receptor PDFDocument46 pagesToll-Like Receptor PDFHeru HanantoNo ratings yet
- Review: Toll-Like Receptors and The Control of ImmunityDocument23 pagesReview: Toll-Like Receptors and The Control of ImmunityIrma NeyraNo ratings yet
- Watts 2008Document3 pagesWatts 2008Ana Julia TicchiNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
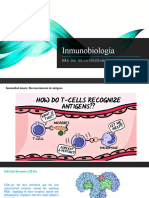
- InmunobiologíaDocument37 pagesInmunobiologíaSoledad Lagunes CastroNo ratings yet
- Viruses: Protective Role of Toll-Like Receptor 3-Induced Type I Interferon in Murine Coronavirus Infection of MacrophagesDocument23 pagesViruses: Protective Role of Toll-Like Receptor 3-Induced Type I Interferon in Murine Coronavirus Infection of MacrophagesRitha PratiwiNo ratings yet
- Review Article: Holding The Inflammatory System in Check: Tlrs and Their Targeted Therapy in AsthmaDocument9 pagesReview Article: Holding The Inflammatory System in Check: Tlrs and Their Targeted Therapy in AsthmaArianRizkiAmaliaNo ratings yet
- MicrobialDerived Tolllike Receptor Agonism in Cancer Treatment and ProgressionCancersDocument20 pagesMicrobialDerived Tolllike Receptor Agonism in Cancer Treatment and ProgressionCancersKajelcha FikaduNo ratings yet
- Gmbar ImunDocument10 pagesGmbar ImunVera FitrianaNo ratings yet
- Receptores Toll-Like Como Un Escape Mecanismo de Defensa Del HuéspedDocument10 pagesReceptores Toll-Like Como Un Escape Mecanismo de Defensa Del HuéspedKArdo23No ratings yet
- Innate Immune Functions of Astrocytes Are Dependent Upon Tumor Necrosis Factor-AlphaDocument15 pagesInnate Immune Functions of Astrocytes Are Dependent Upon Tumor Necrosis Factor-AlphaCony GSNo ratings yet
- The AIM2 Inflammasome,, Sensor of Pathogens and CellularDocument53 pagesThe AIM2 Inflammasome,, Sensor of Pathogens and CellularRadwan AlzidanNo ratings yet
- The Immunopathogenesis of Sepsis: InsightDocument7 pagesThe Immunopathogenesis of Sepsis: InsightMedranoReyesLuisinNo ratings yet
- The Immunopathogenesis of Sepsis: InsightDocument7 pagesThe Immunopathogenesis of Sepsis: InsightnadyaNo ratings yet
- Intracellular Proteins Involved in Inflammation and ApoptosisDocument12 pagesIntracellular Proteins Involved in Inflammation and ApoptosisJUAREZ GONZALEZ EDWARD HASSANNo ratings yet
- Gorp Et Al-2014-FEBS JournalDocument15 pagesGorp Et Al-2014-FEBS JournalTirta ArifNo ratings yet
- Critical Review: Signaling Through Toll-Like Receptor 4 and Mast Cell-Dependent Innate Immunity ResponsesDocument8 pagesCritical Review: Signaling Through Toll-Like Receptor 4 and Mast Cell-Dependent Innate Immunity ResponsesLara SantosNo ratings yet
- Dengue Virus and The Host Innate Immune ResponseDocument6 pagesDengue Virus and The Host Innate Immune ResponseAzalia MacedoNo ratings yet
- An Overview of The Non-Canonical Inflammasome 2020Document13 pagesAn Overview of The Non-Canonical Inflammasome 2020Muhammad AthamnaNo ratings yet
- Inmunidad InnataDocument7 pagesInmunidad InnataWilfrido Jose Barrios AgamezNo ratings yet
- Innate 2005 AbDocument93 pagesInnate 2005 AbAnonymous wVgEAr6No ratings yet
- YuykkDocument2 pagesYuykkImroz ShakilNo ratings yet
- Review Article: Modulation of Immune-Inflammatory Responses in Abdominal Aortic Aneurysm: Emerging Molecular TargetsDocument15 pagesReview Article: Modulation of Immune-Inflammatory Responses in Abdominal Aortic Aneurysm: Emerging Molecular Targetsyenny handayani sihiteNo ratings yet
- Toward Targeting in Ammasomes: Insights Into Their Regulation and ActivationDocument13 pagesToward Targeting in Ammasomes: Insights Into Their Regulation and ActivationRin ChanNo ratings yet
- Immunity and Inflammation in Health and Disease: Emerging Roles of Nutraceuticals and Functional Foods in Immune SupportFrom EverandImmunity and Inflammation in Health and Disease: Emerging Roles of Nutraceuticals and Functional Foods in Immune SupportShampa ChatterjeeNo ratings yet
- Metadata of The Chapter That Will Be Visualized in SpringerlinkDocument17 pagesMetadata of The Chapter That Will Be Visualized in Springerlinknicky_kelly_3No ratings yet
- 2nd LifeDocument7 pages2nd Lifevinit99No ratings yet
- Avatars and Arrows in The BrainDocument11 pagesAvatars and Arrows in The Brainnicky_kelly_3No ratings yet
- Andrew Joron The Chicago Surrealist GroupDocument12 pagesAndrew Joron The Chicago Surrealist Groupnicky_kelly_3No ratings yet
- Hematological Findings and Complications of COVID-19: AffiliationsDocument32 pagesHematological Findings and Complications of COVID-19: Affiliationsnicky_kelly_3No ratings yet
- Rising Inequality and Stagnation in The US EconomyDocument13 pagesRising Inequality and Stagnation in The US Economynicky_kelly_3No ratings yet
- Discovering Language Variations in Whats PDFDocument14 pagesDiscovering Language Variations in Whats PDFnicky_kelly_3No ratings yet
- Should Poetry Be Ethical or Otherwise PDFDocument21 pagesShould Poetry Be Ethical or Otherwise PDFnicky_kelly_3No ratings yet
- Deleuze3053-Masochism Coldness and CrueltyDocument23 pagesDeleuze3053-Masochism Coldness and Crueltyxianbeer100% (2)
- 2015-Journal of Thrombosis and Haemostasis PDFDocument997 pages2015-Journal of Thrombosis and Haemostasis PDFnikospapaioannouNo ratings yet
- Pathophysiology Septic ShockDocument26 pagesPathophysiology Septic ShockTinea Sycillia100% (1)
- Veterinary Immunology and Immunopathology: Research PaperDocument8 pagesVeterinary Immunology and Immunopathology: Research PaperLaura Nataly VillamarinNo ratings yet
- Stenotrophomona Maltophilia y Bulkorelia Cepacia. Mnadell 2009Document8 pagesStenotrophomona Maltophilia y Bulkorelia Cepacia. Mnadell 2009Laura López Del Castillo LalydelcaNo ratings yet
- Artículo 2 - Fisiopatología de La PielonefritisDocument17 pagesArtículo 2 - Fisiopatología de La PielonefritisCAROLINA CHUMACERO BERMEONo ratings yet
- Molecules: Anti-Depressive Effectiveness of Baicalin in Vitro and in VivoDocument13 pagesMolecules: Anti-Depressive Effectiveness of Baicalin in Vitro and in VivoMylena SilvaNo ratings yet
- Fisiopatología de La Meningitis BacterianaDocument7 pagesFisiopatología de La Meningitis BacterianaGiuliano AlbizziNo ratings yet
- Role of Synthetic Proteins and Peptides in AnimalsDocument7 pagesRole of Synthetic Proteins and Peptides in AnimalsroniNo ratings yet
- Toxicology Letters: Sandra Murphy, Robert Ellis-Hutchings, Lavorgie Finch, Stefanie Welz, Karin WienchDocument7 pagesToxicology Letters: Sandra Murphy, Robert Ellis-Hutchings, Lavorgie Finch, Stefanie Welz, Karin WienchBjdjNo ratings yet
- Once The Barrier Is Breached How Are Pathogens Detected and Dealt With?Document64 pagesOnce The Barrier Is Breached How Are Pathogens Detected and Dealt With?Seyeon Clare LeeNo ratings yet
- RP105 and Its Role On PeriodontitisDocument4 pagesRP105 and Its Role On PeriodontitisVijayalaxmi KokatnurNo ratings yet
- Durruty, Et Al (2019) Pathogenesis of Type 2 DiabetesDocument19 pagesDurruty, Et Al (2019) Pathogenesis of Type 2 Diabetestantry puspitasariNo ratings yet
- A Hydrophila PathogenesityDocument66 pagesA Hydrophila PathogenesityDeepak Ranjan SahooNo ratings yet
- (Hyperbaric Oxygen Therapy (Hbot) in Case of Mumps-Induced Sudden Deafness PediatricDocument11 pages(Hyperbaric Oxygen Therapy (Hbot) in Case of Mumps-Induced Sudden Deafness PediatricerickNo ratings yet
- LDN Information (2!19!17 Update)Document18 pagesLDN Information (2!19!17 Update)bktango100% (1)
- CH 03Document16 pagesCH 03TahmidNo ratings yet
- Acetabular Revision Surgery in MajorDocument223 pagesAcetabular Revision Surgery in MajorWilson A. Quintero100% (2)
- Review Article: Mechanisms of Bone Resorption in PeriodontitisDocument11 pagesReview Article: Mechanisms of Bone Resorption in PeriodontitisBetrik Sefyana MangiriNo ratings yet
- Syphilis Recognition Description and DiagnosisDocument138 pagesSyphilis Recognition Description and DiagnosisCelta Gómez TrejoNo ratings yet
- Normal 5caf880433466Document24 pagesNormal 5caf880433466HossanaDaphneLosiusNo ratings yet
- Autoimmunity Reviews: Maximiliano Javier Jiménez-Dalmaroni, M. Eric Gerswhin, Iannis E. AdamopoulosDocument8 pagesAutoimmunity Reviews: Maximiliano Javier Jiménez-Dalmaroni, M. Eric Gerswhin, Iannis E. AdamopoulosTeodor CabelNo ratings yet
- Complement System Part II: Role in ImmunityDocument26 pagesComplement System Part II: Role in ImmunityAnca LunguNo ratings yet
- csb0001011 Full PDFDocument30 pagescsb0001011 Full PDFNate AlveyNo ratings yet
- Adnan EROL, MD. Professor of Internal Medicine EPDH For The Disorders of Energy Metabolism Silivri-Istanbul, TurkeyDocument8 pagesAdnan EROL, MD. Professor of Internal Medicine EPDH For The Disorders of Energy Metabolism Silivri-Istanbul, TurkeyLoisNo ratings yet
- Shinjita Das M.D., Rachel Reynolds M.D. (Auth.), Joshua Zeichner (Eds.) - Acneiform Eruptions in Dermatology - A Differential Diagnosis-Springer-Verlag New York (2014) PDFDocument405 pagesShinjita Das M.D., Rachel Reynolds M.D. (Auth.), Joshua Zeichner (Eds.) - Acneiform Eruptions in Dermatology - A Differential Diagnosis-Springer-Verlag New York (2014) PDFMaryNo ratings yet
- Circulating Heat Shock Protein 60 Levels Are Elevated in HIV Patients and Are Reduced by Anti-Retroviral TherapyDocument8 pagesCirculating Heat Shock Protein 60 Levels Are Elevated in HIV Patients and Are Reduced by Anti-Retroviral TherapyAmalia WidowatiNo ratings yet
- Chapter 42 - Sepsis, Septic Shock, and Multiple Organ FailureDocument15 pagesChapter 42 - Sepsis, Septic Shock, and Multiple Organ FailureWorrawalun FuktongNo ratings yet
- Antiphospholipid Syndrome - From Pathogenesis To Novel Immunomodulatory TherapiesDocument6 pagesAntiphospholipid Syndrome - From Pathogenesis To Novel Immunomodulatory TherapiesardirshajiNo ratings yet
- Alrouji 2018Document47 pagesAlrouji 2018farah maulida martaNo ratings yet
- Neonatal Sepsis: Alicia Pacala, RN BSN Alverno CollegeDocument50 pagesNeonatal Sepsis: Alicia Pacala, RN BSN Alverno CollegeGaladariNo ratings yet