You might also like
- UGA Stratigraphy Lab: Acetate PeelsDocument1 pageUGA Stratigraphy Lab: Acetate PeelsJul JulNo ratings yet
- Newsletter2003 PDFDocument14 pagesNewsletter2003 PDFJul JulNo ratings yet
- European Association of Vertebrate PalaeontologistsDocument4 pagesEuropean Association of Vertebrate PalaeontologistsJul JulNo ratings yet
- Paleontological Type Specimens: Catalogue of Invertebrate and VertebrateDocument214 pagesPaleontological Type Specimens: Catalogue of Invertebrate and VertebrateJul JulNo ratings yet
- Araujo Et Al 2009 - Técnicas de Replicado de Un EstegosaurioDocument24 pagesAraujo Et Al 2009 - Técnicas de Replicado de Un EstegosaurioJul JulNo ratings yet
- Sedighi-Gilani Et Al 2016 - Relationship of Vibro-Mechanical Properties and Microstructure of Wood and Varnish InterfaceDocument12 pagesSedighi-Gilani Et Al 2016 - Relationship of Vibro-Mechanical Properties and Microstructure of Wood and Varnish InterfaceJul JulNo ratings yet
- Buksnowitza & Teischinger 2006 - Resonance Wood - Evaluation and Prediction of Violin Makers' Quality-Grading PDFDocument12 pagesBuksnowitza & Teischinger 2006 - Resonance Wood - Evaluation and Prediction of Violin Makers' Quality-Grading PDFJul JulNo ratings yet
- Tufail Et Al 2017 - Termal Effect On Mechanical Properties of Limestone... BORRAR LUEGODocument12 pagesTufail Et Al 2017 - Termal Effect On Mechanical Properties of Limestone... BORRAR LUEGOJul JulNo ratings yet
- Yavuz Et Al 2010 - Thermal Effect On The Physical Properties of Carbonate Rocks-AnnotatedDocument10 pagesYavuz Et Al 2010 - Thermal Effect On The Physical Properties of Carbonate Rocks-AnnotatedJul JulNo ratings yet
- Pundir Et Al 2009 - Estimation of Age Based On Tooth Cementum Annulations Using Three Different Microscopic MethodsDocument6 pagesPundir Et Al 2009 - Estimation of Age Based On Tooth Cementum Annulations Using Three Different Microscopic MethodsJul JulNo ratings yet
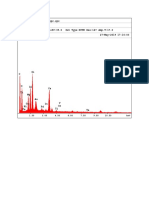
- EDAX Spectrum 1Document2 pagesEDAX Spectrum 1Jul JulNo ratings yet
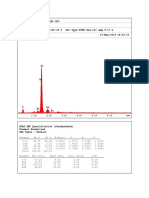
- EDAX Spectrum 8Document1 pageEDAX Spectrum 8Jul JulNo ratings yet
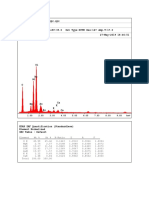
- EDAX Spectrum 9Document2 pagesEDAX Spectrum 9Jul JulNo ratings yet
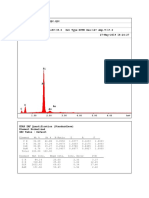
- EDAX Spectrum 6Document1 pageEDAX Spectrum 6Jul JulNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Dental Distress Syndrome QuantifiedDocument37 pagesThe Dental Distress Syndrome QuantifiedDentalbay GrNo ratings yet
- Marshal Cavendish Science - Activity Book With Answer - Stage 4 - C02Document17 pagesMarshal Cavendish Science - Activity Book With Answer - Stage 4 - C02Syed JunaidNo ratings yet
- Blue Boxes SummaryDocument16 pagesBlue Boxes Summaryridin007No ratings yet
- Cervical Spine Compensation in Adolescent Idiopathic ScoliosisDocument5 pagesCervical Spine Compensation in Adolescent Idiopathic Scoliosisshivaprasad kolurNo ratings yet
- Second Periodical Test in ScienceDocument2 pagesSecond Periodical Test in ScienceLorraine lee100% (1)
- Invertebrates 3-4 Unit Guide PDFDocument8 pagesInvertebrates 3-4 Unit Guide PDFFilm & Animation IndiaNo ratings yet
- Assessment For Massage TherapistDocument656 pagesAssessment For Massage Therapistbb_ul100% (7)
- How Bout Them Bones!: Background KnowledgeDocument2 pagesHow Bout Them Bones!: Background KnowledgeVinujah SukumaranNo ratings yet
- Spine Injury: Bassey, A E Dep'T of Orthopaedic & Trauma Surgery Uath, AbujaDocument38 pagesSpine Injury: Bassey, A E Dep'T of Orthopaedic & Trauma Surgery Uath, AbujagamalramadiNo ratings yet
- Musculoskeltal ProbsDocument40 pagesMusculoskeltal ProbsmilayosoresNo ratings yet
- MCQs in OrthopaedicsDocument32 pagesMCQs in OrthopaedicsRakesh KumarNo ratings yet
- Ferret OrthopedicsDocument27 pagesFerret OrthopedicsTayná P DobnerNo ratings yet
- Ruptured and Herniated Disc AnimationDocument4 pagesRuptured and Herniated Disc Animationgaby yepesNo ratings yet
- An Essay On Yoga and HealthDocument2 pagesAn Essay On Yoga and HealthGentleStrength100% (1)
- David Berceli Webinar Notes, Trauma Releasing Exercises NotesDocument23 pagesDavid Berceli Webinar Notes, Trauma Releasing Exercises NotesAnonymous F4KmWCH100% (11)
- Ebook Hadzics Peripheral Nerve Blocks and Anatomy For Ultrasound Guided Regional Anesthesia PDF Full Chapter PDFDocument67 pagesEbook Hadzics Peripheral Nerve Blocks and Anatomy For Ultrasound Guided Regional Anesthesia PDF Full Chapter PDFdonald.mortensen170100% (26)
- Skeletal System - Study MaterialDocument4 pagesSkeletal System - Study MaterialNabeel UndreNo ratings yet
- Exposure Chart: Lumbar Spine Medium Large SmallDocument5 pagesExposure Chart: Lumbar Spine Medium Large Smallkhawar mukhtarNo ratings yet
- Clinical Features and Diagnosis of Neoplastic Epidural Spinal Cord Compression - UpToDateDocument19 pagesClinical Features and Diagnosis of Neoplastic Epidural Spinal Cord Compression - UpToDatePrincess Jeanne Roque GairanodNo ratings yet
- Assessment of The Thorax and LungsDocument108 pagesAssessment of The Thorax and LungsJohn Martin Casihan100% (3)
- Bpes2 SemDocument95 pagesBpes2 Semnishthabanga1525No ratings yet
- Lennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)Document395 pagesLennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)αργυ αργυ100% (1)
- C 06 Bradford II Faunal AnalysisDocument14 pagesC 06 Bradford II Faunal AnalysisCraig ChartierNo ratings yet
- Dca CPT CodesDocument1 pageDca CPT CodesMonstrozeroNo ratings yet
- Campbell's Operative OrthopaedicsDocument5 pagesCampbell's Operative OrthopaedicsShazada Khan50% (4)
- A Field Guide To Joint Disease in Archaelogy - J. Rogers and T. Waldron OCR ReduitDocument66 pagesA Field Guide To Joint Disease in Archaelogy - J. Rogers and T. Waldron OCR ReduitCeilidh Lerwick83% (6)
- Positional TractionDocument3 pagesPositional Tractionicrs_aamir100% (1)
- Script Back SafetyDocument11 pagesScript Back SafetyEsnedaNo ratings yet
- NURSING CASE STUDY: Brain Tumor Grade III "ANAPLASTIC ASTROCYTOMA"Document69 pagesNURSING CASE STUDY: Brain Tumor Grade III "ANAPLASTIC ASTROCYTOMA"EJ Cubero, R☤N100% (27)
- Spinal Orthoses: Type Examples Motion Restriction Indications NotesDocument2 pagesSpinal Orthoses: Type Examples Motion Restriction Indications NotesBD BesorioNo ratings yet