You might also like
- Larvicidal Activity of Celosin A, A Novel Compound Isolated From Celosia Argentea LinnDocument6 pagesLarvicidal Activity of Celosin A, A Novel Compound Isolated From Celosia Argentea LinnInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Screening of Antimicrobial and Antioxidant Secondary Metabolites From Endophytic Fungi Isolated From Wheat (Triticum Durum)Document9 pagesScreening of Antimicrobial and Antioxidant Secondary Metabolites From Endophytic Fungi Isolated From Wheat (Triticum Durum)Hiranda WildayaniNo ratings yet
- Biocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaDocument9 pagesBiocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaMaria BatoolNo ratings yet
- Fungal endophytes inhibit major pathogens of groundnutDocument12 pagesFungal endophytes inhibit major pathogens of groundnutcuong hoNo ratings yet
- Management of Bacterial Leaf Blight Disease in Rice With Endophytic BacteriaDocument18 pagesManagement of Bacterial Leaf Blight Disease in Rice With Endophytic BacteriaArbi Iman Bahtiar NobizNo ratings yet
- Antifungal Activity Pak BotanyDocument8 pagesAntifungal Activity Pak Botanywaheed akramNo ratings yet
- Fajifer, 2022Document6 pagesFajifer, 2022florentinamelania84No ratings yet
- Abo-State Et Al., 2015Document7 pagesAbo-State Et Al., 2015Jordana KalineNo ratings yet
- 5 PDFDocument8 pages5 PDFRobertNo ratings yet
- The Effectiveness of Using Metarhizium Anisopliae,: Submitted byDocument11 pagesThe Effectiveness of Using Metarhizium Anisopliae,: Submitted byMunch AlonzoNo ratings yet
- Funaria Hygrometrica Extracts With Activity Against Plant Pathogenic Fungi Alternaria SpeciesDocument8 pagesFunaria Hygrometrica Extracts With Activity Against Plant Pathogenic Fungi Alternaria SpeciesESSENCE - International Journal for Environmental Rehabilitation and ConservaionNo ratings yet
- Fungi Isolated From Insects in Strawberry Crops As Potential Biological Control DuponcheliaDocument9 pagesFungi Isolated From Insects in Strawberry Crops As Potential Biological Control DuponcheliaJan TenienteNo ratings yet
- 2019 21 3 209 212Document4 pages2019 21 3 209 212Shailendra RajanNo ratings yet
- Paper MicroDocument10 pagesPaper MicroWilliam EliezerNo ratings yet
- tmpBA15 TMPDocument12 pagestmpBA15 TMPFrontiersNo ratings yet
- Tmpa936 TMPDocument12 pagesTmpa936 TMPFrontiersNo ratings yet
- Uji BiokimiaDocument10 pagesUji BiokimiaDandi SaputraNo ratings yet
- Sphaerica and Oscillatoria AgardhiiDocument17 pagesSphaerica and Oscillatoria AgardhiiMhemeydha Luphe YudhaNo ratings yet
- Volatile Compounds Inhibit Anthracnose PathogenDocument7 pagesVolatile Compounds Inhibit Anthracnose PathogenKieu Viet AnhNo ratings yet
- Insecticidal Effect of Dittrichia Viscosa Lyophilized Epicuticular MaterialDocument6 pagesInsecticidal Effect of Dittrichia Viscosa Lyophilized Epicuticular MaterialIlija MileticNo ratings yet
- Environment Friendly Antibacterial Activity of Water Chestnut FruitsDocument9 pagesEnvironment Friendly Antibacterial Activity of Water Chestnut FruitsAnowar RazvyNo ratings yet
- Benelli 2015Document12 pagesBenelli 2015Gustavo AndresNo ratings yet
- B Cereus 1Document11 pagesB Cereus 1Natalia DuqueNo ratings yet
- Ebadollahi 2017Document6 pagesEbadollahi 2017Abdenour AdjaoudNo ratings yet
- Binod 2007Document8 pagesBinod 2007yassen badawyNo ratings yet
- VitaletalDocument6 pagesVitaletalElgie JimenezNo ratings yet
- Antifungal Volatile Organic Compounds From The Endophyte Nodulisporium SP GS4d2II1aDocument18 pagesAntifungal Volatile Organic Compounds From The Endophyte Nodulisporium SP GS4d2II1aCarmen Yuliana GutierrezNo ratings yet
- JPAM Vol 15 Issue1 P 232-239Document8 pagesJPAM Vol 15 Issue1 P 232-239Jefri Nur HidayatNo ratings yet
- Effect of Potential of Beauveria Bassiana and Metarrhizium Sp. To Control The Plutella Xylostella Pest in South KalimanatanDocument8 pagesEffect of Potential of Beauveria Bassiana and Metarrhizium Sp. To Control The Plutella Xylostella Pest in South KalimanatanOpenaccess Research paperNo ratings yet
- 539 31-38 FinalDocument8 pages539 31-38 FinalDavidNo ratings yet
- Screening Antimicrobial Activity Cephalopod Ink Against PathogensDocument9 pagesScreening Antimicrobial Activity Cephalopod Ink Against PathogensNadzirulNo ratings yet
- Virulence of Four Beauveria Bassiana (Balsamo) (Asc., Hypocreales) Isolates On Rose Saw Y, Arge Rosae Under Laboratory ConditionDocument5 pagesVirulence of Four Beauveria Bassiana (Balsamo) (Asc., Hypocreales) Isolates On Rose Saw Y, Arge Rosae Under Laboratory ConditionShellGarciaNo ratings yet
- Antifungal Activity of Macrofungi Extracts Against Phytopathogenic Fungal StrainsDocument10 pagesAntifungal Activity of Macrofungi Extracts Against Phytopathogenic Fungal StrainsDaniel MerelNo ratings yet
- Nicotiana GlaucaDocument7 pagesNicotiana GlaucaNicolas Ignacio AllendeNo ratings yet
- AjenjoDocument9 pagesAjenjovidalNo ratings yet
- 1 s2.0 S0167880916304820 MainDocument9 pages1 s2.0 S0167880916304820 MainQuevedodi IqdNo ratings yet
- Plant extracts inhibit yam rot fungiDocument5 pagesPlant extracts inhibit yam rot fungiFrancis SurveyorNo ratings yet
- Capsicum Annuum: Antibacterial Activities of Endophytic Fungi From L. (Siling Labuyo) Leaves and FruitsDocument10 pagesCapsicum Annuum: Antibacterial Activities of Endophytic Fungi From L. (Siling Labuyo) Leaves and FruitsMeynard Angelo M. JavierNo ratings yet
- Article in Press: Isolation and Identification of An Antibacterial Compound From Diplotaxis Harra (Forssk.) BoissDocument7 pagesArticle in Press: Isolation and Identification of An Antibacterial Compound From Diplotaxis Harra (Forssk.) BoissIpah PahriahNo ratings yet
- s41938-017-0010-3 q1Document12 pagess41938-017-0010-3 q1Tammy CevallosNo ratings yet
- 589 PDF 3Document3 pages589 PDF 3Mustafizur Rahman AviNo ratings yet
- In-Vitro Inhibitory Indices of Selected Fungal Isolates Against Mycotoxin FungiDocument8 pagesIn-Vitro Inhibitory Indices of Selected Fungal Isolates Against Mycotoxin FungiOpenaccess Research paperNo ratings yet
- Applsci 09 02239Document12 pagesApplsci 09 02239Panagiotis EliopoulosNo ratings yet
- Inhibitory efficacy of some agrochemicals on mycelial growth of Phytophthora cinnamomi isolated from heart-rot disease of pineapple (Ananas cosmosus (L.) Merr.)Document5 pagesInhibitory efficacy of some agrochemicals on mycelial growth of Phytophthora cinnamomi isolated from heart-rot disease of pineapple (Ananas cosmosus (L.) Merr.)Openaccess Research paperNo ratings yet
- Jurnal Mikro 2Document5 pagesJurnal Mikro 2Juju JuntakNo ratings yet
- Isolation and Identification of Phytopathogenic Bacteria in Vegetable Crops in West Africa (Côte D'ivoire)Document11 pagesIsolation and Identification of Phytopathogenic Bacteria in Vegetable Crops in West Africa (Côte D'ivoire)dawit gNo ratings yet
- Effects of Neem Insecticides on Cauliflower PestDocument7 pagesEffects of Neem Insecticides on Cauliflower PestDuvánE.DueñasLópezNo ratings yet
- Impact of Diazotrophic Bacteria On Germination and Growth of Tomato, With Bio-Control Effect, Isolated From Algerian SoilDocument7 pagesImpact of Diazotrophic Bacteria On Germination and Growth of Tomato, With Bio-Control Effect, Isolated From Algerian SoilsaidaNo ratings yet
- PR2 GROUP5. PaynalDocument13 pagesPR2 GROUP5. PaynalLale KezNo ratings yet
- Evaluating Oviposition Deterrent Larvicidal and ReDocument14 pagesEvaluating Oviposition Deterrent Larvicidal and RejopatnacarioNo ratings yet
- ANti BiotikDocument7 pagesANti BiotikRidho Dhe HolmesNo ratings yet
- ANTIBACTERIAL ACTIVITIESDocument4 pagesANTIBACTERIAL ACTIVITIESMD MuntaharNo ratings yet
- Theobroma Cacao PhytophthoraDocument10 pagesTheobroma Cacao PhytophthoraOpenaccess Research paperNo ratings yet
- 38.july 2013 1373366194 cf2bb 23Document6 pages38.july 2013 1373366194 cf2bb 23davikNo ratings yet
- Alimudin GMR22Document10 pagesAlimudin GMR22Rosyida Ismi (Rosyi)No ratings yet
- Beauveria Bassiana Metarhizium AnisopliaeDocument8 pagesBeauveria Bassiana Metarhizium AnisopliaeYuliastuti YasminNo ratings yet
- Biocontrol of Tomato Bacterial Wilt by Foliar Spray Application of A Novel Strain of Endophytic Bacillus Sp.Document11 pagesBiocontrol of Tomato Bacterial Wilt by Foliar Spray Application of A Novel Strain of Endophytic Bacillus Sp.Moni Becerra WongNo ratings yet
- 5 PaperDocument6 pages5 PaperDr. Nilesh Baburao JawalkarNo ratings yet
- Plant Disease Management Strategies for Sustainable Agriculture through Traditional and Modern ApproachesFrom EverandPlant Disease Management Strategies for Sustainable Agriculture through Traditional and Modern ApproachesImran Ul HaqNo ratings yet
- IJHST Volume 4 Issue 2 Pages 239-250Document12 pagesIJHST Volume 4 Issue 2 Pages 239-250Cedrus CedrusNo ratings yet
- Selective: Medium For Isolation of Species DematiaceousDocument4 pagesSelective: Medium For Isolation of Species DematiaceousCedrus CedrusNo ratings yet
- 1029 - (Identification - and - Pathogenicity - Assessment - of - Fus... )Document11 pages1029 - (Identification - and - Pathogenicity - Assessment - of - Fus... )Cedrus CedrusNo ratings yet
- 413 Ijar-19602Document7 pages413 Ijar-19602Cedrus CedrusNo ratings yet
- Isolation and Identification of Seed Associated Fungi of WheatDocument4 pagesIsolation and Identification of Seed Associated Fungi of WheatCedrus CedrusNo ratings yet
- Dervis 2010Document15 pagesDervis 2010Cedrus CedrusNo ratings yet
- Thanassoulopoulos 1993Document4 pagesThanassoulopoulos 1993Cedrus CedrusNo ratings yet
- Verticillium Wilt in Tennessee Valley CottonDocument7 pagesVerticillium Wilt in Tennessee Valley CottonCedrus CedrusNo ratings yet
- Occurrence of Defoliating Verticillium dahliae in Tunisian OliveDocument6 pagesOccurrence of Defoliating Verticillium dahliae in Tunisian OliveCedrus CedrusNo ratings yet
- A Brief Overview of Seed Priming Benefits in Tomato: Original PaperDocument9 pagesA Brief Overview of Seed Priming Benefits in Tomato: Original PaperCedrus CedrusNo ratings yet
- Verticillium Wilts: A Review of Control Options and ExternalitiesDocument12 pagesVerticillium Wilts: A Review of Control Options and ExternalitiesCedrus CedrusNo ratings yet
- Effect of Seed Priming On Seed Quality of Tomato (Solanum Lycoperscium L.)Document4 pagesEffect of Seed Priming On Seed Quality of Tomato (Solanum Lycoperscium L.)Cedrus CedrusNo ratings yet
- Rodriguez 2004Document12 pagesRodriguez 2004Cedrus CedrusNo ratings yet
- Han Et Al. 2016Document10 pagesHan Et Al. 2016Cedrus CedrusNo ratings yet
- Technical and Physical A Spects of Energy Saving in GreenhousesDocument1 pageTechnical and Physical A Spects of Energy Saving in GreenhousesCedrus CedrusNo ratings yet
- Forest Soil Influence on Ectomycorrhizal DevelopmentDocument12 pagesForest Soil Influence on Ectomycorrhizal DevelopmentCedrus CedrusNo ratings yet
- Acta Hort 03Document45 pagesActa Hort 03Cedrus CedrusNo ratings yet
- 9 Basant Chhetri FinalDocument6 pages9 Basant Chhetri FinalCedrus CedrusNo ratings yet
- IOBC2016 AnnouncementCfADocument2 pagesIOBC2016 AnnouncementCfACedrus CedrusNo ratings yet
- Komatsud65ex 16dozerbulldozerservicerepairmanualsn80001andup 200727063646Document26 pagesKomatsud65ex 16dozerbulldozerservicerepairmanualsn80001andup 200727063646juan santa cruzNo ratings yet
- Starting A Business Candle Making 2009Document2 pagesStarting A Business Candle Making 2009Carlo Fabros Junio100% (4)
- Translation of Japanese Onomatopoeia Into Swedish (With Focus On Lexicalization)Document20 pagesTranslation of Japanese Onomatopoeia Into Swedish (With Focus On Lexicalization)Aldandy OckadeyaNo ratings yet
- Administracion Una Perspectiva Global Y Empresarial Resumen Por CapitulosDocument7 pagesAdministracion Una Perspectiva Global Y Empresarial Resumen Por Capitulosafmqqaepfaqbah100% (1)
- More Than Moore: by M. Mitchell WaldropDocument4 pagesMore Than Moore: by M. Mitchell WaldropJuanjo ThepresisNo ratings yet
- Visualization BenchmarkingDocument15 pagesVisualization BenchmarkingRanjith S100% (1)
- Lateral capacity of pile in clayDocument10 pagesLateral capacity of pile in clayGeetha MaNo ratings yet
- Emilio Aguinaldo CollegeDocument1 pageEmilio Aguinaldo CollegeRakeshKumar1987No ratings yet
- QP P1 APR 2023Document16 pagesQP P1 APR 2023Gil legaspiNo ratings yet
- Business EnvironmentDocument12 pagesBusiness EnvironmentAbhinav GuptaNo ratings yet
- ClientsDocument7 pagesClientsLiane PanahacNo ratings yet
- POPULARITY OF CREDIT CARDS ISSUED BY DIFFERENT BANKSDocument25 pagesPOPULARITY OF CREDIT CARDS ISSUED BY DIFFERENT BANKSNaveed Karim Baksh75% (8)
- Solutions: Spheres, Cones and CylindersDocument13 pagesSolutions: Spheres, Cones and CylindersKeri-ann MillarNo ratings yet
- Getting the Most from Cattle Manure: Proper Application Rates and PracticesDocument4 pagesGetting the Most from Cattle Manure: Proper Application Rates and PracticesRamNocturnalNo ratings yet
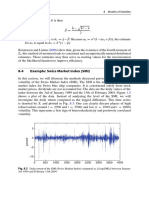
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- MATHEMATICAL ECONOMICSDocument54 pagesMATHEMATICAL ECONOMICSCities Normah0% (1)
- Presepsi Khalayak Terhadap Program Acara Televise Reality Show "Jika Aku Menjadi" Di Trans TVDocument128 pagesPresepsi Khalayak Terhadap Program Acara Televise Reality Show "Jika Aku Menjadi" Di Trans TVAngga DianNo ratings yet
- English Extra Conversation Club International Women's DayDocument2 pagesEnglish Extra Conversation Club International Women's Dayevagloria11No ratings yet
- 3 - Content - Introduction To Java, JVM, JDK PDFDocument8 pages3 - Content - Introduction To Java, JVM, JDK PDFAnonymous zdY202lgZYNo ratings yet
- Hanwha Engineering & Construction - Brochure - enDocument48 pagesHanwha Engineering & Construction - Brochure - enAnthony GeorgeNo ratings yet
- District Wise List of Colleges Under The Juridiction of MRSPTU BathindaDocument13 pagesDistrict Wise List of Colleges Under The Juridiction of MRSPTU BathindaGurpreet SandhuNo ratings yet
- Operating Manual: Please Read These Operating Instructions Before Using Your FreedomchairDocument24 pagesOperating Manual: Please Read These Operating Instructions Before Using Your FreedomchairNETHYA SHARMANo ratings yet
- Muv PDFDocument6 pagesMuv PDFDenisse PxndithxNo ratings yet
- 13Document47 pages13Rohan TirmakheNo ratings yet
- Quick Reference - HVAC (Part-1) : DECEMBER 1, 2019Document18 pagesQuick Reference - HVAC (Part-1) : DECEMBER 1, 2019shrawan kumarNo ratings yet
- X RayDocument16 pagesX RayMedical Physics2124No ratings yet
- Galambos 1986Document18 pagesGalambos 1986gcoNo ratings yet
- Project Report On Evaporative CoolerDocument12 pagesProject Report On Evaporative Coolersourabh singh tomerNo ratings yet
- AIOUDocument2 pagesAIOUHoorabwaseemNo ratings yet
- PedigreesDocument5 pagesPedigreestpn72hjg88No ratings yet