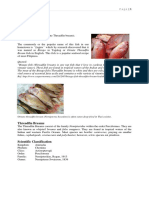
You might also like
- Gold V Essex CCDocument11 pagesGold V Essex CCZACHARIAH MANKIRNo ratings yet
- Stranger Things Episode Script 2 05 Chapter Five Dig DugDocument67 pagesStranger Things Episode Script 2 05 Chapter Five Dig Dugyonas123No ratings yet
- Is College For Everyone - RevisedDocument5 pagesIs College For Everyone - Revisedapi-295480043No ratings yet
- #2 Chapter IIDocument17 pages#2 Chapter IIAngelyn Bantilo SerronaNo ratings yet
- Chapter 18Document10 pagesChapter 18Aeriel Venice VergaraNo ratings yet
- Saltwater Crocodile PDFDocument12 pagesSaltwater Crocodile PDFfkhunc070% (1)
- Download Shells Of The World A Natural History A Guide To Every Family 10 Harasewych all chapterDocument77 pagesDownload Shells Of The World A Natural History A Guide To Every Family 10 Harasewych all chapterlessie.stewart123100% (5)
- A Review On Status and Conservation of Saltwater Crocodile (Crocodylus Porosus) in IndiaDocument9 pagesA Review On Status and Conservation of Saltwater Crocodile (Crocodylus Porosus) in IndiaMuhammad Fernanda DanuartaNo ratings yet
- Chordata NotesDocument5 pagesChordata NotesAbhishek Isaac MathewNo ratings yet
- Motani2009 Article TheEvolutionOfMarineReptilesDocument12 pagesMotani2009 Article TheEvolutionOfMarineReptilesFlorentin MiuNo ratings yet
- EchinodermDocument63 pagesEchinodermMarilyn Buac OgocNo ratings yet
- Klasifikasi VertebrataDocument13 pagesKlasifikasi VertebrataFony Pellokila SinlaeNo ratings yet
- 4 Chapter IIDocument18 pages4 Chapter IIAngelyn Bantilo SerronaNo ratings yet
- Cetaceans: What They Are and ExamplesDocument3 pagesCetaceans: What They Are and ExamplesDavid Contreras LopezNo ratings yet
- Short Notes: Bird Study June 2009Document5 pagesShort Notes: Bird Study June 2009Ivan VujkovicNo ratings yet
- First Semester 2020/2021Document53 pagesFirst Semester 2020/2021low luffecussNo ratings yet
- Lopezlopez 2012Document9 pagesLopezlopez 2012Fadhli LatuconsinaNo ratings yet
- Current Final EchinodermsDocument23 pagesCurrent Final EchinodermsMikaela Rome Bigay100% (1)
- 2021 MolluskDocument14 pages2021 MolluskAkash GautamNo ratings yet
- Marine Biology Irs - Marine MammalsDocument11 pagesMarine Biology Irs - Marine MammalsMaureen HelmNo ratings yet
- Did You Know ThatDocument10 pagesDid You Know ThatCheska Dean VergaraNo ratings yet
- Molluscs in Mangroves: A Case Study: Prof. A. Shanmugam and Mrs. S. VairamaniDocument12 pagesMolluscs in Mangroves: A Case Study: Prof. A. Shanmugam and Mrs. S. VairamaniUsdiyantoNo ratings yet
- Bearez 1998 - FishingPoisonSeaEnvironmentSalango PDFDocument6 pagesBearez 1998 - FishingPoisonSeaEnvironmentSalango PDFNayelitzin J. CanoNo ratings yet
- Occurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaDocument10 pagesOccurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaRizky RamaNo ratings yet
- Killer Whale Classification and CharacteristicsDocument6 pagesKiller Whale Classification and CharacteristicsFase BakNo ratings yet
- Animal Diversity - The Kingdom Animalia 1. Phylum Porifera - SpongesDocument6 pagesAnimal Diversity - The Kingdom Animalia 1. Phylum Porifera - SpongesSamantha Dasmariñas MagaanNo ratings yet
- 039-054 HoganDocument16 pages039-054 HoganJonathan Villamil RodríguezNo ratings yet
- Abjna 2 6 897 901Document5 pagesAbjna 2 6 897 901zakiNo ratings yet
- Ecology of Coral Reef Fishes (L.E. Hallacher)Document59 pagesEcology of Coral Reef Fishes (L.E. Hallacher)speedphobic07100% (1)
- The First Vertebrates, Jawless Fishes, The Agnathans: 2.1 OstracodermsDocument22 pagesThe First Vertebrates, Jawless Fishes, The Agnathans: 2.1 OstracodermsAlejandro Tepoz TelloNo ratings yet
- Squid: Scientific Name AppearanceDocument23 pagesSquid: Scientific Name AppearanceMageswari SelvamNo ratings yet
- Ross 2000Document40 pagesRoss 2000nicole delgadoNo ratings yet
- Cap. 02 - General Anatomy and Physiology of ReptilesDocument23 pagesCap. 02 - General Anatomy and Physiology of ReptilesNailson JúniorNo ratings yet
- LagawDocument3 pagesLagawMarjon Velasco Dela Cerna IINo ratings yet
- Klasifikasi Dan Identifikasi Penyu - Id.enDocument5 pagesKlasifikasi Dan Identifikasi Penyu - Id.enPrangko SembilantujuhNo ratings yet
- Presentation2 130406234902 Phpapp02Document18 pagesPresentation2 130406234902 Phpapp02mimilNo ratings yet
- Browse Search Login: Article Fast Facts Media Additional InfoDocument4 pagesBrowse Search Login: Article Fast Facts Media Additional Infococoh43288No ratings yet
- Wiki SnailDocument1 pageWiki Snailglh00No ratings yet
- Fish Predation by Semi-Aquatic Spiders: A Global Pattern: Martin Nyffeler, Bradley J. PuseyDocument21 pagesFish Predation by Semi-Aquatic Spiders: A Global Pattern: Martin Nyffeler, Bradley J. PuseyAndreas SchmitzNo ratings yet
- Rommel Etal 2006Document22 pagesRommel Etal 2006Erven DerenNo ratings yet
- MOLLUSCA 3 GastropodaDocument14 pagesMOLLUSCA 3 GastropodaCARLOSNo ratings yet
- White PaperDocument8 pagesWhite Paperapi-549082571No ratings yet
- Protect parrotfish to protect reefsDocument2 pagesProtect parrotfish to protect reefsHaliliNo ratings yet
- Bony FishesDocument70 pagesBony FishesJigar PatelNo ratings yet
- The European Shark GuideDocument23 pagesThe European Shark GuideOur Compass75% (4)
- Reptiles OriginEvolutionDocument11 pagesReptiles OriginEvolutionSeptianisa UlfalahNo ratings yet
- Animalia 2Document63 pagesAnimalia 2Rida IshanurNo ratings yet
- C or Are: Echinoderms Class OphiuroidsDocument10 pagesC or Are: Echinoderms Class Ophiuroidsprashanth1989sNo ratings yet
- Reporters: Maun, Maja Mariella Gonzales, Alessy Joy Cortez, KateDocument34 pagesReporters: Maun, Maja Mariella Gonzales, Alessy Joy Cortez, KateKate CortezNo ratings yet
- ProjectDocument5 pagesProjectAbid ShaikhNo ratings yet
- Phylum Annelida (Segmented Worms) : College of Science Biological Sciences DepartmentDocument22 pagesPhylum Annelida (Segmented Worms) : College of Science Biological Sciences DepartmentPrateek JainNo ratings yet
- Literature Review of Asteroidea Sea StarsDocument11 pagesLiterature Review of Asteroidea Sea StarsSett SambajonNo ratings yet
- Crocodile in The Indus Civilisation ParpolaDocument58 pagesCrocodile in The Indus Civilisation ParpolaKarlNo ratings yet
- Caribbean Seagrasses As A Food Source For The Emerald Neritid Smaragdia ViridisDocument7 pagesCaribbean Seagrasses As A Food Source For The Emerald Neritid Smaragdia ViridispulpopulpoNo ratings yet
- Reptiles OriginEvolutionDocument11 pagesReptiles OriginEvolutionPascoal MiguelNo ratings yet
- Pseudocoelomates Phyla OutlineDocument8 pagesPseudocoelomates Phyla Outlinesteanne100% (1)
- Univerzitet U Beogradu Biološki Fakultet: Razviće VodozemacaDocument25 pagesUniverzitet U Beogradu Biološki Fakultet: Razviće VodozemacaAndjelka ZivkovicNo ratings yet
- Fish Are Gill-Bearing Aquatic Craniate Animals That Lack Limbs With DigitsDocument24 pagesFish Are Gill-Bearing Aquatic Craniate Animals That Lack Limbs With Digitssai calderNo ratings yet
- Life and Role of Primitive Vertebrate: The Guide Lecture: Dr. Safrida, S. PD., M. SiDocument23 pagesLife and Role of Primitive Vertebrate: The Guide Lecture: Dr. Safrida, S. PD., M. SiGina anggrianaNo ratings yet
- Diversity of Fish: by SpeciesDocument9 pagesDiversity of Fish: by SpeciessamiNo ratings yet
- Sea Animals WKSDocument7 pagesSea Animals WKSBianca CardozoNo ratings yet
- Deep Thinkers: Inside the Minds of Whales, Dolphins, and PorpoisesFrom EverandDeep Thinkers: Inside the Minds of Whales, Dolphins, and PorpoisesNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 3 5 PDFDocument3 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 3 5 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 2 4 PDFDocument3 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 2 4 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 1 2 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 1 2 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 11 12 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 11 12 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 6 7 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 6 7 PDFharryNo ratings yet
- The Domesticated Donkey: I - Species Characteristics: October 2012Document2 pagesThe Domesticated Donkey: I - Species Characteristics: October 2012harryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 15 16 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 15 16 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 8 9 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 8 9 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 13 14 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 13 14 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 11 12 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 11 12 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 3 5 PDFDocument3 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 3 5 PDFharryNo ratings yet
- Ballscrew Product Specifications 7 8Document2 pagesBallscrew Product Specifications 7 8harryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 6 7 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 6 7 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 13 14 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 13 14 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 8 9 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 8 9 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 15 16 PDFDocument2 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 15 16 PDFharryNo ratings yet
- A2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 2 4 PDFDocument3 pagesA2 27 2012 Ylmazetal TheDomesticatedDonkeyIEconomicImportanceUncommonUsageetc 2 4 PDFharryNo ratings yet
- Center Distance (Permissible Rotation Speed) : Fig. 3 Screw Shaft Mounting Method: Fixed - FixedDocument3 pagesCenter Distance (Permissible Rotation Speed) : Fig. 3 Screw Shaft Mounting Method: Fixed - FixedharryNo ratings yet
- Ballscrew Product Specifications 11 12Document2 pagesBallscrew Product Specifications 11 12harryNo ratings yet
- Ballscrew Product Specifications 3 4Document2 pagesBallscrew Product Specifications 3 4harryNo ratings yet
- Ballscrew Product Specifications 19 20Document2 pagesBallscrew Product Specifications 19 20harryNo ratings yet
- Ballscrew Product Specifications 13 14Document2 pagesBallscrew Product Specifications 13 14harryNo ratings yet
- Ballscrew Product Specifications 26 27Document2 pagesBallscrew Product Specifications 26 27harryNo ratings yet
- Ballscrew Product Specifications 5 6Document2 pagesBallscrew Product Specifications 5 6harryNo ratings yet
- Ballscrew Product Specifications 9 10Document2 pagesBallscrew Product Specifications 9 10harryNo ratings yet
- Ballscrew Product Specifications 15 16Document2 pagesBallscrew Product Specifications 15 16harryNo ratings yet
- Heat Generation Test: Unused Ball Ball After Traveling Unused Ball Ball After TravelingDocument2 pagesHeat Generation Test: Unused Ball Ball After Traveling Unused Ball Ball After TravelingharryNo ratings yet
- Ballscrew Product Specifications 17 18Document2 pagesBallscrew Product Specifications 17 18harryNo ratings yet
- Ballscrew Product Specifications 24 25Document2 pagesBallscrew Product Specifications 24 25harryNo ratings yet
- Ballscrew Product Specifications 1 2Document2 pagesBallscrew Product Specifications 1 2harryNo ratings yet
- F K Up This Book Sample PDFDocument38 pagesF K Up This Book Sample PDFBlack & White PublishingNo ratings yet
- Tugas B.INGGRIS ALANDocument4 pagesTugas B.INGGRIS ALANAlan GunawanNo ratings yet
- Album China RayaDocument12 pagesAlbum China Rayaapitmelodi23No ratings yet
- Ch01 Standard Methods and PractisesDocument44 pagesCh01 Standard Methods and PractisesUsman FarooqNo ratings yet
- 515 AMA Must DO QuestionsDocument217 pages515 AMA Must DO QuestionsVenkataRajuNo ratings yet
- Philips NTRX505Document28 pagesPhilips NTRX505EulerMartinsDeMelloNo ratings yet
- DSE2541 Data SheetDocument2 pagesDSE2541 Data SheetBERANGER DAVESNE DJOMALIA SIEWENo ratings yet
- The Kano Model: How To Delight Your CustomersDocument16 pagesThe Kano Model: How To Delight Your CustomersRoen RamonalNo ratings yet
- Ish300 Chapter 3Document20 pagesIsh300 Chapter 3Amanina AyuniNo ratings yet
- NCM-111B - Concept Paper. Impact of COVID-19Document10 pagesNCM-111B - Concept Paper. Impact of COVID-19Ivy VillalobosNo ratings yet
- Thời gian làm bài: 90 phút (không kể thời gian giao đề)Document8 pagesThời gian làm bài: 90 phút (không kể thời gian giao đề)Nguyễn KiênNo ratings yet
- June 10Document16 pagesJune 10rogeliodmngNo ratings yet
- 21st Century Theories of EducationDocument53 pages21st Century Theories of Educationdaffodil198100% (1)
- Alat Studio Dan KomunikasiDocument14 pagesAlat Studio Dan Komunikasiraymon akbarNo ratings yet
- JAWABAN UTS Bahasa Inggris II Keperawatan Dewi YuniarDocument3 pagesJAWABAN UTS Bahasa Inggris II Keperawatan Dewi Yuniarovan maysandyNo ratings yet
- TELENGANA BUSINESS DIRECTORY PENEL D-PanelDocument65 pagesTELENGANA BUSINESS DIRECTORY PENEL D-Panelamandeep100% (4)
- News TIA Portal V15 and V15 1 enDocument43 pagesNews TIA Portal V15 and V15 1 enjohanNo ratings yet
- Sheet-Pan Salmon and Broccoli With Sesame and Ginger: by Lidey HeuckDocument2 pagesSheet-Pan Salmon and Broccoli With Sesame and Ginger: by Lidey HeuckllawNo ratings yet
- FisikaDocument46 pagesFisikaNurol Hifzi Putri RizkiNo ratings yet
- Survey of Accounting 6th Edition Warren Solutions ManualDocument17 pagesSurvey of Accounting 6th Edition Warren Solutions Manualdevinsmithddsfzmioybeqr100% (19)
- 2.3 Stabilized MaterialDocument11 pages2.3 Stabilized MaterialMamush LetaNo ratings yet
- Part of Speech QuestionsDocument4 pagesPart of Speech QuestionsNi Luh Ari Kusumawati0% (1)
- 03 IoT Technical Sales Training Industrial Wireless Deep DiveDocument35 pages03 IoT Technical Sales Training Industrial Wireless Deep Divechindi.comNo ratings yet
- Marantz sr4200 Service PDFDocument29 pagesMarantz sr4200 Service PDFAnonymous KSedwANo ratings yet
- EthicsDocument10 pagesEthicsEssi Chan100% (4)
- Read Me 22222222222222Document2 pagesRead Me 22222222222222sancakemreNo ratings yet
- Medicard Phil Inc. vs. CIRDocument2 pagesMedicard Phil Inc. vs. CIRhigoremso giensdksNo ratings yet