You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Delirium en Paciente OncologicoDocument12 pagesDelirium en Paciente OncologicoCatalina RodriguezNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Delirium: Continuum Lifelong Learning Neurol 2010 16 (2) :120-134Document15 pagesDelirium: Continuum Lifelong Learning Neurol 2010 16 (2) :120-134Maria Isabel Montañez RestrepoNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Developmental Cognitive NeuroscienceDocument10 pagesDevelopmental Cognitive NeuroscienceMatheus Paes PeçanhaNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Sex Differences in The Developing Brain As ADocument12 pagesSex Differences in The Developing Brain As AMaria Isabel Montañez RestrepoNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Neuroimmunology and Neuroepigenetics in The Establishment of Sex Differences in The BrainDocument32 pagesNeuroimmunology and Neuroepigenetics in The Establishment of Sex Differences in The BrainMaria Isabel Montañez RestrepoNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Sexual Identity Development Among Gay, Lesbian, and Bisexual PDFDocument24 pagesSexual Identity Development Among Gay, Lesbian, and Bisexual PDFMaria Isabel Montañez RestrepoNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Sex Differences of Brain and Their Implications For Personalized TherapyDocument14 pagesSex Differences of Brain and Their Implications For Personalized TherapyMaria Isabel Montañez RestrepoNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Homosexual Orientation. The Search For A Biological Basis.: References ReprintsDocument5 pagesHomosexual Orientation. The Search For A Biological Basis.: References ReprintsMaria Isabel Montañez RestrepoNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- 18721116: Riluzole in Psychiatry A Systematic Review of The LiteratureDocument17 pages18721116: Riluzole in Psychiatry A Systematic Review of The LiteratureMaria Isabel Montañez RestrepoNo ratings yet
- Pedophilia-Themed Obsessive-Compulsive Disorder: Assessment, Differential Diagnosis, and Treatment With Exposure and Response PreventionDocument14 pagesPedophilia-Themed Obsessive-Compulsive Disorder: Assessment, Differential Diagnosis, and Treatment With Exposure and Response PreventionMaria Isabel Montañez RestrepoNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Characteristics of Unacceptable/taboo Thoughts in Obsessive - Compulsive DisorderDocument8 pagesThe Characteristics of Unacceptable/taboo Thoughts in Obsessive - Compulsive DisorderMaria Isabel Montañez RestrepoNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Neurology of Acquired Pedophilia: NeurocaseDocument13 pagesThe Neurology of Acquired Pedophilia: NeurocaseMaria Isabel Montañez RestrepoNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Epidemiology of Obsessive-Compulsive Disorder: ICD-IO CriteriaDocument5 pagesEpidemiology of Obsessive-Compulsive Disorder: ICD-IO CriteriaMaria Isabel Montañez RestrepoNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- AnsiedadDocument33 pagesAnsiedadMaria Isabel Montañez RestrepoNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Breedlove SM Watson NV The Minds Machine Foundation of BrainDocument602 pagesBreedlove SM Watson NV The Minds Machine Foundation of BrainElena Chebotayeva100% (2)
- Psychosomatics: Digestive System: Miguel Agustin ST Francisco, RPM, Mpsy CandDocument45 pagesPsychosomatics: Digestive System: Miguel Agustin ST Francisco, RPM, Mpsy CandRalph Christian ZorillaNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Biological Effects of Androgen On Male and FemaleDocument22 pagesBiological Effects of Androgen On Male and FemaleMarie Elexis RevillezaNo ratings yet
- Insights New111Document13 pagesInsights New111cassidy conchaNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Sarah Richardson - Plasticity and Programming (2017)Document24 pagesSarah Richardson - Plasticity and Programming (2017)SofíaRBNo ratings yet
- Sex Determination and Sex Differentiation in FishDocument174 pagesSex Determination and Sex Differentiation in Fishtrunghieu85No ratings yet
- Evidence Supporting The Biological Nature of Gender IdentityDocument6 pagesEvidence Supporting The Biological Nature of Gender IdentityMorgan Atwood100% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Ambiguous GenitaliaDocument47 pagesAmbiguous GenitaliaDrvsowmya Lakshmi100% (1)
- Bao Swaab 2011Document13 pagesBao Swaab 2011BeatrizCamposNo ratings yet
- Sex DifferentiationDocument59 pagesSex DifferentiationAndreoli CorreiaNo ratings yet
- 1975 Book IntersexualityInTheAnimalKingdDocument464 pages1975 Book IntersexualityInTheAnimalKingdDiego RodriguesNo ratings yet
- Hormones and Behavior: Jill E. Schneider, Jeremy M. Brozek, Erin Keen-RhinehartDocument16 pagesHormones and Behavior: Jill E. Schneider, Jeremy M. Brozek, Erin Keen-RhinehartKerin ArdyNo ratings yet
- Hormones & BehaviorDocument34 pagesHormones & BehavioravanthikaNo ratings yet
- Evaluation of The Infant With Atypical Genitalia (Disorder of Sex Development) - UpToDateDocument40 pagesEvaluation of The Infant With Atypical Genitalia (Disorder of Sex Development) - UpToDateKumara GuruNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Original PDF An Introduction To Behavioral Endocrinology 5th Edition PDFDocument41 pagesOriginal PDF An Introduction To Behavioral Endocrinology 5th Edition PDFrichard.logsdon482100% (38)
- Behavioral Neurobiology Developmental Psychobiology and Behavioral EcologyDocument469 pagesBehavioral Neurobiology Developmental Psychobiology and Behavioral Ecologyluis recaldeNo ratings yet
- Dick Swaab - Sexual Differentiation of The Human Brain in Relation To Gender Identity and Sexual OrientationDocument24 pagesDick Swaab - Sexual Differentiation of The Human Brain in Relation To Gender Identity and Sexual OrientationWalyson de SousaNo ratings yet
- Harry Benjamin Syndrome As An Intersex ConditionDocument19 pagesHarry Benjamin Syndrome As An Intersex ConditionVien TanjungNo ratings yet
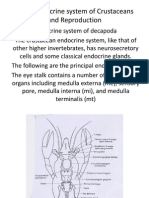
- Neuroendocrine of CrustaceaDocument29 pagesNeuroendocrine of CrustaceaAnirudh Acharya100% (1)
- Sex Determination and DifferentiationDocument17 pagesSex Determination and DifferentiationAaron LeeNo ratings yet
- The Biology of Human Psychosexual Differentiation: Louis GoorenDocument13 pagesThe Biology of Human Psychosexual Differentiation: Louis GoorenDavid EspinolaNo ratings yet
- Chapter 15. Sex Determination and Differentiation Barry Sinervo©1997-2004Document21 pagesChapter 15. Sex Determination and Differentiation Barry Sinervo©1997-2004Helen Diaz-PaezNo ratings yet
- Human Sexuality Self Society and Culture 1st Edition Herdt Solutions ManualDocument23 pagesHuman Sexuality Self Society and Culture 1st Edition Herdt Solutions Manualdianbandelethek6xo100% (19)
- 2010 - Sex Differences in The Brain, Behavior, and Neuropsychiatric DisordersDocument16 pages2010 - Sex Differences in The Brain, Behavior, and Neuropsychiatric DisordersDiego Alejandro Londoño CorreaNo ratings yet
- Julie A Greenberg - Intersexuality and The Law Why Sex MattersDocument180 pagesJulie A Greenberg - Intersexuality and The Law Why Sex MattersLaura Belli100% (1)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)