You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Titan GelDocument6 pagesTitan GelVladi Salas100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Logarithmic Functions Week 7Document20 pagesLogarithmic Functions Week 7gadraNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Nema MG10 PDFDocument27 pagesNema MG10 PDFManuel Antonio Santos VargasNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Nurtured Womb e BookDocument22 pagesNurtured Womb e BookSteph's Desserts100% (1)
- Famous Bombers of The Second World War - 1st SeriesDocument142 pagesFamous Bombers of The Second World War - 1st Seriesgunfighter29100% (1)
- Pitman SolutionDocument190 pagesPitman SolutionBon Siranart50% (2)
- Foundation Design LectureDocument59 pagesFoundation Design LectureJamaica MarambaNo ratings yet
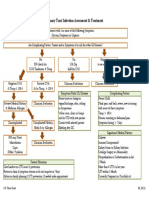
- Inclusion Criteria:: Urinary Tract Infection Care MapDocument9 pagesInclusion Criteria:: Urinary Tract Infection Care MapMoh. H.AlObaidyNo ratings yet
- Infection Prevention-Related Needs Assessment Needs AssessmentDocument10 pagesInfection Prevention-Related Needs Assessment Needs AssessmentMoh. H.AlObaidyNo ratings yet
- Nursing Care 2Document1 pageNursing Care 2Moh. H.AlObaidyNo ratings yet
- Ambulance Operations) ThisDocument2 pagesAmbulance Operations) ThisMoh. H.AlObaidyNo ratings yet
- Abdominal Radiography: Procedure OverviewDocument6 pagesAbdominal Radiography: Procedure OverviewMoh. H.AlObaidyNo ratings yet
- Notice - Appeal Process List of Appeal Panel (Final 12.1.24)Document13 pagesNotice - Appeal Process List of Appeal Panel (Final 12.1.24)FyBerri InkNo ratings yet
- ESM-4810A1 Energy Storage Module User ManualDocument31 pagesESM-4810A1 Energy Storage Module User ManualOscar SosaNo ratings yet
- 3TRIGODocument5 pages3TRIGOCielo Pabilan ObemioNo ratings yet
- HACH LANGE Amino Acid F Reagent Powder (2353255)Document6 pagesHACH LANGE Amino Acid F Reagent Powder (2353255)kerem__22No ratings yet
- Operating Manual CSDPR-V2-200-NDocument19 pagesOperating Manual CSDPR-V2-200-NJohnTPNo ratings yet
- CFD Analysis of Flow Through Compressor CascadeDocument10 pagesCFD Analysis of Flow Through Compressor CascadeKhalid KhalilNo ratings yet
- AssignmentDocument13 pagesAssignmentSwakshar DebNo ratings yet
- Battery Installation ProcedureDocument5 pagesBattery Installation ProceduresantoshkumarNo ratings yet
- Chapter 01Document16 pagesChapter 01deepak_baidNo ratings yet
- Introduction To Kalman FilterDocument4 pagesIntroduction To Kalman FilterArghya MukherjeeNo ratings yet
- Etl 213-1208.10 enDocument1 pageEtl 213-1208.10 enhossamNo ratings yet
- Accessories 162-USDocument20 pagesAccessories 162-USعايد التعزيNo ratings yet
- List ToolingDocument10 pagesList Toolingyohanes adi saputroNo ratings yet
- Setting Times of ConcreteDocument3 pagesSetting Times of ConcreteP DhanunjayaNo ratings yet
- Feasibility Study On The Seaweed Kappaphycus Alvarezii Cultivation Site in Indari Waters ofDocument9 pagesFeasibility Study On The Seaweed Kappaphycus Alvarezii Cultivation Site in Indari Waters ofUsman MadubunNo ratings yet
- Water Chemistry - An Introduction To The Chemistry of Natural and Engineered Aquatic Systems-Páginas-483-492Document10 pagesWater Chemistry - An Introduction To The Chemistry of Natural and Engineered Aquatic Systems-Páginas-483-492jhonier guevaraNo ratings yet
- English 3 Avicenna Graded Test 1Document11 pagesEnglish 3 Avicenna Graded Test 1Mohd FarisNo ratings yet
- SweetenersDocument23 pagesSweetenersNur AfifahNo ratings yet
- Fo Transfer ProcedureDocument8 pagesFo Transfer ProcedureadityasahayNo ratings yet
- Sandvik Saf 31803 Tube and Pipe, Seamless: DatasheetDocument9 pagesSandvik Saf 31803 Tube and Pipe, Seamless: DatasheetPaul NeedhamNo ratings yet
- Defect Origin (J) Phase Requirement Analysis Design Coding Unit TestingDocument87 pagesDefect Origin (J) Phase Requirement Analysis Design Coding Unit Testingashish_jain_csNo ratings yet
- Sample Dewa Inspection CommentsDocument2 pagesSample Dewa Inspection Commentsrmtaqui100% (1)
- QIAGEN Price List 2017Document62 pagesQIAGEN Price List 2017Dayakar Padmavathi Boddupally80% (5)