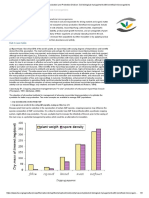
You might also like
- Bacterial Endophytes: Recent Developments and ApplicationsDocument9 pagesBacterial Endophytes: Recent Developments and ApplicationsmaczweiNo ratings yet
- Fevo 10 833389Document12 pagesFevo 10 833389clingoNo ratings yet
- tmpD493 TMPDocument11 pagestmpD493 TMPFrontiersNo ratings yet
- Mycorrhiza in Various Habitats Including The Mechanisms For Survival in Ecosystems and Enhanced Crop Production.Document8 pagesMycorrhiza in Various Habitats Including The Mechanisms For Survival in Ecosystems and Enhanced Crop Production.NaiomiNo ratings yet
- Fmicb 08 01874Document10 pagesFmicb 08 01874lhagwaa BayanjargalNo ratings yet
- Jordan Et Al-2000-Weed ResearchDocument15 pagesJordan Et Al-2000-Weed ResearchMixtli López OrozcoNo ratings yet
- Teotia Et Al. 2017 Movilización de Nutrientes Por HM - 1128377282Document19 pagesTeotia Et Al. 2017 Movilización de Nutrientes Por HM - 1128377282fin03205No ratings yet
- Review On Role of Vam Fungi in Crop Plant-Soil System: Ameeta Sharma & Savita YadavDocument8 pagesReview On Role of Vam Fungi in Crop Plant-Soil System: Ameeta Sharma & Savita YadavTJPRC PublicationsNo ratings yet
- Nodulation, Growth and Yield Response of Five Cowpea (VignaDocument12 pagesNodulation, Growth and Yield Response of Five Cowpea (VignaPremier PublishersNo ratings yet
- Whipps Et Al, 2008Document12 pagesWhipps Et Al, 2008joyeeta8No ratings yet
- Supervivencia y Crecimiento de Fraxinus Uhdei, Inoculado en Vivero, en Cárcavas de AcrisolesDocument8 pagesSupervivencia y Crecimiento de Fraxinus Uhdei, Inoculado en Vivero, en Cárcavas de AcrisolesLuisDomínguezNo ratings yet
- TMP 65Document12 pagesTMP 65FrontiersNo ratings yet
- J Funeco 2021 101087Document12 pagesJ Funeco 2021 101087Raymundo Lopez NNo ratings yet
- Agriculture, Ecosystems and Environment: Tim Dieko Tter, Sabine Wamser, Volkmar Wolters, Klaus BirkhoferDocument5 pagesAgriculture, Ecosystems and Environment: Tim Dieko Tter, Sabine Wamser, Volkmar Wolters, Klaus BirkhoferDHBNo ratings yet
- Soil Biology & Biochemistry: S. Hortal, F. Bastida, J.L. Moreno, C. Armas, C. García, F.I. PugnaireDocument10 pagesSoil Biology & Biochemistry: S. Hortal, F. Bastida, J.L. Moreno, C. Armas, C. García, F.I. PugnaireKássia RamosNo ratings yet
- 1-S2.0-S0378112722004455-Main HONGOS EUROPADocument13 pages1-S2.0-S0378112722004455-Main HONGOS EUROPAEdward Condori PumaNo ratings yet
- Soil Biology & Biochemistry: Xiaoke Zhang, Howard Ferris, Jeffrey Mitchell, Wenju LiangDocument8 pagesSoil Biology & Biochemistry: Xiaoke Zhang, Howard Ferris, Jeffrey Mitchell, Wenju Liangbriologo2No ratings yet
- Jurnal Internasional The Effect of ManureDocument11 pagesJurnal Internasional The Effect of ManureIkram AffandiNo ratings yet
- Plant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsDocument3 pagesPlant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsCyberSquare IndiaNo ratings yet
- Biology Letters - Community Ecology (Bardgett)Document5 pagesBiology Letters - Community Ecology (Bardgett)PololoNo ratings yet
- A Metagenomic-Based, Cross-Seasonal Picture of Fungal CDocument9 pagesA Metagenomic-Based, Cross-Seasonal Picture of Fungal CSoto Agudelo RicardoNo ratings yet
- The Unseen Majority Soil Microbes As Drivers of Plant 1yjo7xbywuDocument15 pagesThe Unseen Majority Soil Microbes As Drivers of Plant 1yjo7xbywuQwerty QwertyNo ratings yet
- Presentation On Fungi SymbiosisDocument19 pagesPresentation On Fungi SymbiosisFemi GajiNo ratings yet
- Ekologi 8Document16 pagesEkologi 8tuti lestariantiNo ratings yet
- Biofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesDocument7 pagesBiofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesIJEAB JournalNo ratings yet
- Jurnal Peran Fungi Dalan EkosistemDocument14 pagesJurnal Peran Fungi Dalan EkosistemEunike Hana GraciaNo ratings yet
- Russian AggDocument7 pagesRussian AggPriyankush Protim BoraNo ratings yet
- 1996 Bever EsporulacaoDocument13 pages1996 Bever EsporulacaoMaristela VolpatoNo ratings yet
- Journey Through Signaling in Arbuscular Mycorrhizal Symbioses.Document13 pagesJourney Through Signaling in Arbuscular Mycorrhizal Symbioses.lourens.darrianNo ratings yet
- Impact of Litter Species Diversity On Decomposition Processes and CommunitiesDocument11 pagesImpact of Litter Species Diversity On Decomposition Processes and CommunitiesFenky SandiNo ratings yet
- TMP 4 CD6Document10 pagesTMP 4 CD6FrontiersNo ratings yet
- MycorhizesDocument9 pagesMycorhizesdoujamaroufNo ratings yet
- Agriculturally Important Fungi: Plant-Microbe Association For Mutual BenefitsDocument20 pagesAgriculturally Important Fungi: Plant-Microbe Association For Mutual BenefitsAnonymous 9xXl8wmBNo ratings yet
- Mastering Ectomycorrhizal Symbiosis: The Impact of CarbohydratesDocument12 pagesMastering Ectomycorrhizal Symbiosis: The Impact of Carbohydratesdatura49No ratings yet
- Application of Organic Fertilizer Changes The Rhizosphere Hg7hqjj4Document15 pagesApplication of Organic Fertilizer Changes The Rhizosphere Hg7hqjj4markgomez451No ratings yet
- Final ManuscriptDocument52 pagesFinal ManuscriptJudea Sevilla DizonNo ratings yet
- Edaphic Invertebrate Macrofauna Associated With Cassava Crop in Different Soil CoveragesDocument8 pagesEdaphic Invertebrate Macrofauna Associated With Cassava Crop in Different Soil CoveragesNathalia FrançaNo ratings yet
- Terpenoids in Plant and Arbuscular Mycorrhiza-Reinforced Defence Against Herbivorous InsectsDocument11 pagesTerpenoids in Plant and Arbuscular Mycorrhiza-Reinforced Defence Against Herbivorous InsectsyogiNo ratings yet
- PhyllosphereDocument38 pagesPhyllosphereRuchi TikarihaNo ratings yet
- Understanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersDocument16 pagesUnderstanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersNancy HutchinsonNo ratings yet
- tmpAEC8 TMPDocument12 pagestmpAEC8 TMPFrontiersNo ratings yet
- Effect of The Combination Rhizophagus Intraradices and 50% of NPK On Maize (Zea Mays L.) Growth and Yield in Central Benin - IJAAR-Vol-16-No-1-p-1-15Document15 pagesEffect of The Combination Rhizophagus Intraradices and 50% of NPK On Maize (Zea Mays L.) Growth and Yield in Central Benin - IJAAR-Vol-16-No-1-p-1-15International Network For Natural SciencesNo ratings yet
- Community Ecology of Ectomycorrhizal Fungi An Advancing Interdisciplinary FieldDocument8 pagesCommunity Ecology of Ectomycorrhizal Fungi An Advancing Interdisciplinary FieldAline FrançaNo ratings yet
- Applied Soil Ecology: Walid Ellouze, Chantal Hamel, Sadok Bouzid, Marc St-ArnaudDocument10 pagesApplied Soil Ecology: Walid Ellouze, Chantal Hamel, Sadok Bouzid, Marc St-ArnaudJorge CorderoNo ratings yet
- Kumar 2017Document48 pagesKumar 2017Wendy BautistaNo ratings yet
- Protistas Do SoloDocument31 pagesProtistas Do Solosalomaofelix576No ratings yet
- Soil Bacterial Taxonomic Diversity Is Critical To Maintaining The PlantDocument7 pagesSoil Bacterial Taxonomic Diversity Is Critical To Maintaining The PlantValkiria Luisa Borsa PiroliNo ratings yet
- Title: Seed Mucilage Interacts With Soil Microbial Community and Physiochemical ProcessesDocument54 pagesTitle: Seed Mucilage Interacts With Soil Microbial Community and Physiochemical Processesben liuNo ratings yet
- Journal of Ecology - 2010 - Scharfy - Invasion of Solidago Gigantea in Contrasting Experimental Plant Communities EffectsDocument10 pagesJournal of Ecology - 2010 - Scharfy - Invasion of Solidago Gigantea in Contrasting Experimental Plant Communities Effectsraizabuenaflor30No ratings yet
- Fpls 04 00165Document7 pagesFpls 04 00165Dicson Sánchez AbadNo ratings yet
- Agronomy 10 01415Document12 pagesAgronomy 10 01415AB LimaNo ratings yet
- Utilization of Arbuscular Micorrhizal Fungi To Control Fusarium Wilt of TomatoesDocument6 pagesUtilization of Arbuscular Micorrhizal Fungi To Control Fusarium Wilt of TomatoeswafiyyatunnufusNo ratings yet
- 1 s2.0 S1470160X21003836 MainDocument11 pages1 s2.0 S1470160X21003836 MainMarcelo Aparecido MarquesNo ratings yet
- CYJS-16-45-85 (Alters Soil Microbial Populations To Facilitate Its Own Growth)Document8 pagesCYJS-16-45-85 (Alters Soil Microbial Populations To Facilitate Its Own Growth)JuanManuelAmaroLuisNo ratings yet
- Effect of Various Crop Production Systems On Population of Rhizosphere Microorganisms in Field Bean CropDocument10 pagesEffect of Various Crop Production Systems On Population of Rhizosphere Microorganisms in Field Bean CropRamesha LiNo ratings yet
- Laprak PTTT Seudah RevisiDocument29 pagesLaprak PTTT Seudah RevisiwafiyyatunnufusNo ratings yet
- The Occurrence of Pesticides in Environment and Current Technologies For Their Remediation and ManagementDocument36 pagesThe Occurrence of Pesticides in Environment and Current Technologies For Their Remediation and ManagementIJAR JOURNALNo ratings yet
- Aggangan-Moon2013 Article TheEffectsOfSoilSterilizationMDocument12 pagesAggangan-Moon2013 Article TheEffectsOfSoilSterilizationMKeren MacielNo ratings yet
- Contreras Cornejo 2017Document9 pagesContreras Cornejo 2017Karen Avalos VelaNo ratings yet
- García-Gómez 2011 TempladoDocument13 pagesGarcía-Gómez 2011 TempladoWendy BautistaNo ratings yet
- O'connell and Bolger 1996Document21 pagesO'connell and Bolger 1996Wendy BautistaNo ratings yet
- Brussaard 1997Document9 pagesBrussaard 1997Wendy BautistaNo ratings yet
- TRicholoma EuropaDocument28 pagesTRicholoma EuropaWendy BautistaNo ratings yet
- Ramaria EspeciesDocument23 pagesRamaria EspeciesWendy BautistaNo ratings yet
- Zinke 2000Document7 pagesZinke 2000Wendy BautistaNo ratings yet
- Ferguson Joly 2002Document9 pagesFerguson Joly 2002Wendy BautistaNo ratings yet
- Timber Forwarding - Physical Soil Properties - Toivio Et Al 2017Document9 pagesTimber Forwarding - Physical Soil Properties - Toivio Et Al 2017Wendy BautistaNo ratings yet
- Sierra Juarez Oax Soils - Alvarez-Arteaga Et Al 2008Document20 pagesSierra Juarez Oax Soils - Alvarez-Arteaga Et Al 2008Wendy BautistaNo ratings yet
- Kumar 2017Document48 pagesKumar 2017Wendy BautistaNo ratings yet
- G ROUNDNUTDocument99 pagesG ROUNDNUTGreen ComputersNo ratings yet
- Biological Nitrogen Fixation: Its Mechanism and Role of Boron in Nitrogen FixationDocument5 pagesBiological Nitrogen Fixation: Its Mechanism and Role of Boron in Nitrogen FixationZorinawmi KhiangteNo ratings yet
- Effect of Vermicompost and Biofertilizers On Yield and Soil PDFDocument3 pagesEffect of Vermicompost and Biofertilizers On Yield and Soil PDFAgricultureASDNo ratings yet
- Types of Bio FertilizersDocument3 pagesTypes of Bio FertilizersKumar ManojNo ratings yet
- Role of Microbes in Organic FarmingDocument13 pagesRole of Microbes in Organic FarmingAvila DCNo ratings yet
- Bio Fertilizer Udyami - Org.inDocument16 pagesBio Fertilizer Udyami - Org.inAndreea DeeaNo ratings yet
- Biofertilizer For Crop Production and Soil Fertility: August 2018Document9 pagesBiofertilizer For Crop Production and Soil Fertility: August 2018GnanakumarNo ratings yet
- Biological Nitrogen FixationDocument24 pagesBiological Nitrogen FixationRam Nivas Ahirwar100% (2)
- Biofertilizer Kenya StandardDocument25 pagesBiofertilizer Kenya StandarddiniindahNo ratings yet
- Bio-FIT Book EN PDFDocument256 pagesBio-FIT Book EN PDFCesar Augusto Colorado RamirezNo ratings yet
- A Ar122eDocument307 pagesA Ar122emongreatNo ratings yet
- C S e Mains Old PaperDocument59 pagesC S e Mains Old PapermoolsinghNo ratings yet
- RS3782 Grade - 12 Sustainable Integrated Nutrient and Pest ManagementDocument84 pagesRS3782 Grade - 12 Sustainable Integrated Nutrient and Pest ManagementAtlantic ChaudharyNo ratings yet
- Chapter2-Soybean Growth andDocument8 pagesChapter2-Soybean Growth andConstantin IancuNo ratings yet
- Biological Process in Soil: Mayrina FirdayatiDocument19 pagesBiological Process in Soil: Mayrina FirdayatiEster MelindaNo ratings yet
- Niir Complete Technology Book On Biofertilizer Organic Farming 2nd Revised EditionDocument12 pagesNiir Complete Technology Book On Biofertilizer Organic Farming 2nd Revised EditionNatalieNo ratings yet
- Nickel Tolerance and Biosorption Potential of Rhizobia Associated With Horse Gram Macrotyloma Uniflorum Lam VerdcDocument8 pagesNickel Tolerance and Biosorption Potential of Rhizobia Associated With Horse Gram Macrotyloma Uniflorum Lam VerdcTajammal Khan DaultanaNo ratings yet
- Concepts of BiofertilizersDocument8 pagesConcepts of BiofertilizersVasu MathuraNo ratings yet
- Effect of Rhizobium Based BiofertilizerDocument10 pagesEffect of Rhizobium Based BiofertilizerMihaela MihaNo ratings yet
- 12 Mineral NutritionDocument5 pages12 Mineral NutritionDe DasNo ratings yet
- 2 18kp1belb1 2020120403355711Document75 pages2 18kp1belb1 2020120403355711RishikeshNo ratings yet
- Of Nitrogen-Fixing Systems: Application in Soil ManagementDocument193 pagesOf Nitrogen-Fixing Systems: Application in Soil Managementjuan cristhianNo ratings yet
- Isolation, Characterization and Purification of Groundnut (Arachis Hypogaea L.)Document10 pagesIsolation, Characterization and Purification of Groundnut (Arachis Hypogaea L.)Andrés SánchezNo ratings yet
- Subject Enrichment Activity Unit Ii Class8Document14 pagesSubject Enrichment Activity Unit Ii Class8rajesh duaNo ratings yet
- Soil Microbiology Ecology and Biochemist PDFDocument13 pagesSoil Microbiology Ecology and Biochemist PDFJuan Jose Granados QuirozNo ratings yet
- Class 7 Holiday Homework TermIDocument6 pagesClass 7 Holiday Homework TermISameeta SarafNo ratings yet
- BioreportDocument8 pagesBioreportAbigail CostalesNo ratings yet
- Directory 2020 ExtensionDocument57 pagesDirectory 2020 ExtensionYared Mesfin FikaduNo ratings yet
- MSC Thesis Updated Today PDFDocument89 pagesMSC Thesis Updated Today PDFDaguale Melaku AyeleNo ratings yet
- Application FormDocument11 pagesApplication FormDarius JacksonNo ratings yet