100% found this document useful (1 vote)
17K views11 pagesVascular Cambium in Botany Assignment
1. The document is an assignment submitted by M Asfand Yaar to Sir Akther on the topic of vascular cambium for a 2nd semester BS Chemistry course at the Govt Muncipal Degree College Faisalabad.
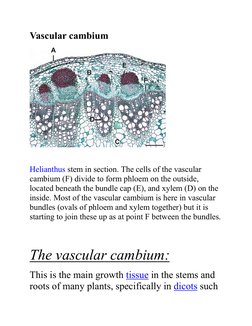
2. The vascular cambium is a layer of meristematic tissue in the stems and roots of vascular plants that produces secondary xylem cells inward and secondary phloem cells outward, allowing for growth in diameter.
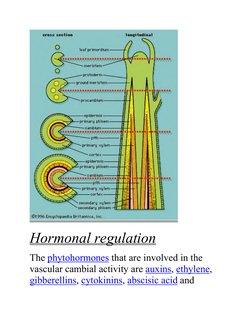
3. The vascular cambium is maintained through a network of hormone signaling including auxins, ethylene, gibberellins, and cytokinins, which regulate processes like cell division, differentiation, and growth.
Uploaded by
Syed Haider Ali Shah SheraziCopyright
© © All Rights Reserved
We take content rights seriously. If you suspect this is your content, claim it here.
Available Formats
Download as PDF, TXT or read online on Scribd
100% found this document useful (1 vote)
17K views11 pagesVascular Cambium in Botany Assignment
1. The document is an assignment submitted by M Asfand Yaar to Sir Akther on the topic of vascular cambium for a 2nd semester BS Chemistry course at the Govt Muncipal Degree College Faisalabad.
2. The vascular cambium is a layer of meristematic tissue in the stems and roots of vascular plants that produces secondary xylem cells inward and secondary phloem cells outward, allowing for growth in diameter.
3. The vascular cambium is maintained through a network of hormone signaling including auxins, ethylene, gibberellins, and cytokinins, which regulate processes like cell division, differentiation, and growth.
Uploaded by
Syed Haider Ali Shah SheraziCopyright
© © All Rights Reserved
We take content rights seriously. If you suspect this is your content, claim it here.
Available Formats
Download as PDF, TXT or read online on Scribd
- Maintenance of cambial meristem: Describes how the vascular cambium is maintained through signal feedback loops and cellular growth.
- Structure and function: Details the structure and physiological role of vascular cambium in plant growth and development.
- Hormonal regulation: Explores the hormonal control of cambial activity focusing on auxins, ethylene, and other hormones.
- See also: Lists additional topics related to vascular cambium for further exploration.
- External links: Provides external links for additional resources related to vascular cambium.
- References: Cites sources and references used for compiling information about vascular cambium.