You might also like
- C9e Answers Active Reading 07Document7 pagesC9e Answers Active Reading 07Daniel DHizzy Hizgilov83% (18)
- Askeland Science and Engineering 7e ISM Chapter 08Document24 pagesAskeland Science and Engineering 7e ISM Chapter 08Ian Gabriel Cañas Fernández100% (4)
- Nelson Biology Answers 1Document2 pagesNelson Biology Answers 1ritaNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture OutlineJohn Westly S. SabueroNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture Outlinehaha_le12No ratings yet
- The Cell MembraneDocument36 pagesThe Cell Membraneanulorance98No ratings yet
- 2 4 Plasma MembraneDocument22 pages2 4 Plasma Membraneıamnıkolaı 4No ratings yet
- Study Exercise Biology 10-12 97.03Document7 pagesStudy Exercise Biology 10-12 97.03Fabiola Vania FeliciaNo ratings yet
- Zoology Lecture 4Document18 pagesZoology Lecture 4Stephen OcheaNo ratings yet
- SOM 201 Cellular MembranesDocument34 pagesSOM 201 Cellular Membraneskxng crockedNo ratings yet
- NSSI Gen Bio 1 Module No Answer KeyDocument43 pagesNSSI Gen Bio 1 Module No Answer Keygian leepsuNo ratings yet
- Fluid Mosaic ModelDocument3 pagesFluid Mosaic Modelmariam manzoorNo ratings yet
- Cell MebraneDocument10 pagesCell MebraneChandra SihombingNo ratings yet
- BIO 315. NotesDocument42 pagesBIO 315. NotesKevin KipropNo ratings yet
- Tissue BiochemistryDocument3 pagesTissue BiochemistryNeelam MehtabNo ratings yet
- Lesson 4: Cell Membrane and Its PermeabilityDocument12 pagesLesson 4: Cell Membrane and Its PermeabilityremshopNo ratings yet
- BIOMEMBRANES Transport MechanismsDocument43 pagesBIOMEMBRANES Transport Mechanismsc3rberussNo ratings yet
- Kuliah MembranDocument61 pagesKuliah Membrannikmah nuur rochmahNo ratings yet
- CSIR Life Science Sample Cell BiologyDocument17 pagesCSIR Life Science Sample Cell Biologygamer zoneNo ratings yet
- Cell MembraneDocument2 pagesCell MembranegrxcieeNo ratings yet
- Membrane Structure and Function ReviewDocument5 pagesMembrane Structure and Function ReviewAnastasiafynnNo ratings yet
- This Learning Competency Is Extracted From The Deped Curriculum GuideDocument6 pagesThis Learning Competency Is Extracted From The Deped Curriculum GuideZuzuNo ratings yet
- 1.3 Membrane StructureDocument6 pages1.3 Membrane StructureMukhammadaminkhon AnvarkhonovNo ratings yet
- Cell and Molecular BiologyDocument2 pagesCell and Molecular BiologyAttaullah36No ratings yet
- Cell Membrane (Plasma Membrane)Document6 pagesCell Membrane (Plasma Membrane)Rica NorcioNo ratings yet
- Cell Membranes and Transport As BiologyDocument41 pagesCell Membranes and Transport As BiologyADEEL AHMADNo ratings yet
- Biologi CHP 6 Sem 1Document81 pagesBiologi CHP 6 Sem 1Aliaa AkbarNo ratings yet
- Plasma Membrane The Big PictureDocument4 pagesPlasma Membrane The Big PictureNicolas Gomez CombellasNo ratings yet
- BIOWEEK5RANESDocument29 pagesBIOWEEK5RANESvrbatitangNo ratings yet
- 5 - Biochemistry - 3rd Year Chem - Biological Membranes and Transports - Chapter-5 - Part 3Document27 pages5 - Biochemistry - 3rd Year Chem - Biological Membranes and Transports - Chapter-5 - Part 3MD. SAJID GHUFRANNo ratings yet
- Plasma Membrane Structure and FunctionsDocument22 pagesPlasma Membrane Structure and FunctionsMian SahibNo ratings yet
- Life at The Edge Concept 7.1: Structure! Cellular Membranes Are Fluid Mosaics of Lipids and ProteinsDocument6 pagesLife at The Edge Concept 7.1: Structure! Cellular Membranes Are Fluid Mosaics of Lipids and ProteinsAisha100% (1)
- The Cell MembraneDocument45 pagesThe Cell MembraneKirito HatakuNo ratings yet
- Crop Science Notes 2 2015Document67 pagesCrop Science Notes 2 2015Alfred Magwede100% (1)
- Chapter 7Document4 pagesChapter 7Astrii LyNo ratings yet
- Local Media2437312247486514219Document29 pagesLocal Media2437312247486514219Olive Vergara MuscaNo ratings yet
- How eukaryotic cells organize into specialized structuresDocument6 pagesHow eukaryotic cells organize into specialized structuresjuan mondaNo ratings yet
- Components of Plasma Membranes: Learning ObjectivesDocument14 pagesComponents of Plasma Membranes: Learning ObjectivesAlthea Junsan BernabeNo ratings yet
- Cell Membrane Structure and FunctionsDocument48 pagesCell Membrane Structure and FunctionsSamNo ratings yet
- Chapter 1doc 3Document4 pagesChapter 1doc 3Enju ChipepeNo ratings yet
- Plasma MembraneDocument18 pagesPlasma MembraneJemuel Bucud LagartoNo ratings yet
- Chapter 2-BacteriaDocument4 pagesChapter 2-BacteriaNGUYEN CONG DUONGNo ratings yet
- DAT Bootcamp Bio Academy: Chapter 2: Cells and OrganellesDocument48 pagesDAT Bootcamp Bio Academy: Chapter 2: Cells and OrganellesEmilyNo ratings yet
- Fluid Mosaic Model Cell Membrane StructureDocument24 pagesFluid Mosaic Model Cell Membrane StructureADEEL AHMADNo ratings yet
- Cells and Genomes MacromoleculesDocument10 pagesCells and Genomes MacromoleculesAkmad SugodNo ratings yet
- Microbiology 2Document25 pagesMicrobiology 2Ba SitNo ratings yet
- Structure and Organization of MembranesDocument20 pagesStructure and Organization of MembranesronojoysenguptaNo ratings yet
- Biomembrane: Selective PermeabilityDocument2 pagesBiomembrane: Selective PermeabilityKrystel LahomNo ratings yet
- Cell BioDocument27 pagesCell BioSumit KumarNo ratings yet
- Struct-Function Cell OrgaDocument30 pagesStruct-Function Cell OrgaSiaw Sing OngNo ratings yet
- Unit 2 ReviewerDocument4 pagesUnit 2 Revieweraaleah moscaNo ratings yet
- Functional and Structural Properties of Natural BiomembranesDocument7 pagesFunctional and Structural Properties of Natural BiomembranesAhsan RazaNo ratings yet
- What Is A MembraneDocument16 pagesWhat Is A MembranesameerNo ratings yet
- 4 - Cell MembranesDocument23 pages4 - Cell Membraneswhether913No ratings yet
- Membrane StructureDocument20 pagesMembrane StructureErnitaNo ratings yet
- LO Week 7Document22 pagesLO Week 7Matthew ChristopherNo ratings yet
- 3 B Cytology-Membrane StructuresDocument19 pages3 B Cytology-Membrane Structuresm.mansoor3377No ratings yet
- Cell Functions & Organelles in 40 CharactersDocument14 pagesCell Functions & Organelles in 40 Charactersamber tariqNo ratings yet
- Cell Biology - DissertationDocument7 pagesCell Biology - Dissertationprerana chakrabortyNo ratings yet
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- Environmental factors leading to major health issuesDocument6 pagesEnvironmental factors leading to major health issuesKrizziane Ivy CuajotorNo ratings yet
- Philippine Games JSTORDocument15 pagesPhilippine Games JSTORKrizziane Ivy CuajotorNo ratings yet
- Cruz Ni MagellanDocument3 pagesCruz Ni MagellanRenz Bryle AlgabreNo ratings yet
- 67-Article Text-118-1-10-20181206Document12 pages67-Article Text-118-1-10-20181206MadelNo ratings yet
- 5 Parlor NG LahiDocument2 pages5 Parlor NG LahiJet Yvan Conejo0% (1)
- EASE STRESS Acrostic (Cuajotor)Document1 pageEASE STRESS Acrostic (Cuajotor)Krizziane Ivy CuajotorNo ratings yet
- Mammary Glands Bio LEC ReportDocument4 pagesMammary Glands Bio LEC ReportKrizziane Ivy CuajotorNo ratings yet
- KadangDocument2 pagesKadangLiemark Gonzaga93% (15)
- Ants NotesDocument7 pagesAnts NotesKrizziane Ivy CuajotorNo ratings yet
- Lecture Outline: Chapter 12 The Cell Cycle Overview: The Key Roles of Cell DivisionDocument12 pagesLecture Outline: Chapter 12 The Cell Cycle Overview: The Key Roles of Cell DivisionKrizziane Ivy CuajotorNo ratings yet
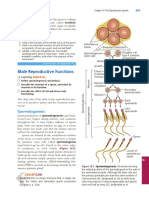
- Male Reproductive Functions: Did You Get It?Document1 pageMale Reproductive Functions: Did You Get It?Krizziane Ivy CuajotorNo ratings yet
- Psych 38 Filipino GamesDocument4 pagesPsych 38 Filipino GamesKrizziane Ivy CuajotorNo ratings yet
- Health Belief ModelDocument2 pagesHealth Belief ModelKrizziane Ivy CuajotorNo ratings yet
- PHASE III (A) - Completion (Wrapping It Up)Document32 pagesPHASE III (A) - Completion (Wrapping It Up)Krizziane Ivy CuajotorNo ratings yet
- Classification of EpitheliaDocument50 pagesClassification of EpitheliaKrizziane Ivy CuajotorNo ratings yet
- Chapter6 - A Tour of The CellDocument20 pagesChapter6 - A Tour of The CellOsannah IrishNo ratings yet
- Chapter13 - Meiosis and Sexual Life CyclesDocument10 pagesChapter13 - Meiosis and Sexual Life CyclesKrizziane Ivy CuajotorNo ratings yet
- Add SurveyDocument1 pageAdd SurveyKrizziane Ivy CuajotorNo ratings yet
- Chapter 1 Exploring Life Overview: Biology's Most Exciting EraDocument14 pagesChapter 1 Exploring Life Overview: Biology's Most Exciting EraKrizziane Ivy CuajotorNo ratings yet
- Gender, Sexuality and the Impact of Divorce in the PhilippinesDocument55 pagesGender, Sexuality and the Impact of Divorce in the PhilippinesKrizziane Ivy CuajotorNo ratings yet
- Amphibians ReportDocument48 pagesAmphibians ReportKrizziane Ivy CuajotorNo ratings yet
- Psych 11 Self-Study Notes (Nov 20, 2017 Monday)Document2 pagesPsych 11 Self-Study Notes (Nov 20, 2017 Monday)Krizziane Ivy CuajotorNo ratings yet
- The History of Plumbing: Krizziane Ivy T. Cuajotor GE7 - Section CDocument20 pagesThe History of Plumbing: Krizziane Ivy T. Cuajotor GE7 - Section CKrizziane Ivy CuajotorNo ratings yet
- BC12 SurveyqDocument2 pagesBC12 SurveyqKrizziane Ivy CuajotorNo ratings yet
- Burner Data SheetDocument15 pagesBurner Data Sheetali_irvNo ratings yet
- Safety CatalogDocument107 pagesSafety CatalogMiguel G V PNo ratings yet
- 26 - Improved Forged Crankshaft Performance Utilizing Deep RollingDocument23 pages26 - Improved Forged Crankshaft Performance Utilizing Deep RollingandypressNo ratings yet
- Photocatalytic Properties of ZrO2 and FeZrO2 SemiconductorsDocument11 pagesPhotocatalytic Properties of ZrO2 and FeZrO2 SemiconductorsBulan Wening R ANo ratings yet
- March 2019 (v2) MS - Paper 5 CIE Biology A-LevelDocument8 pagesMarch 2019 (v2) MS - Paper 5 CIE Biology A-Leveleric sivaneshNo ratings yet
- PDF 13 1Document141 pagesPDF 13 1TsihatesfaNo ratings yet
- An Experimental Study On Strength of Concrete With Silica Fume and Partial Replacement of Cement by Brick PowderDocument5 pagesAn Experimental Study On Strength of Concrete With Silica Fume and Partial Replacement of Cement by Brick PowderInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Design Topic: Introduction To Biology Subject(s) : Biology Grade(s) : 9th Designer(s) : William RutherfordDocument2 pagesDesign Topic: Introduction To Biology Subject(s) : Biology Grade(s) : 9th Designer(s) : William RutherfordWilliam RutherfordNo ratings yet
- 1 s2.0 S0165993621003095 MainDocument11 pages1 s2.0 S0165993621003095 MainSebastian CasadiegoNo ratings yet
- Con Duc To Me TryDocument5 pagesCon Duc To Me TryNareshmurty NareshNo ratings yet
- Magnetic Properties and BH Curve OverviewDocument5 pagesMagnetic Properties and BH Curve Overviewbenitogaldos19gmail.comNo ratings yet
- 5.1 Titremetric Analysis - Direct, Indirect and Residual TitrationDocument2 pages5.1 Titremetric Analysis - Direct, Indirect and Residual TitrationSister RislyNo ratings yet
- Alfa Laval tj40g - Product Leaflet Ese03366enDocument4 pagesAlfa Laval tj40g - Product Leaflet Ese03366enSanket SabaleNo ratings yet
- 2004 Modeling Wood Gasification in A Countercurrent Fixed Bed ReactorDocument14 pages2004 Modeling Wood Gasification in A Countercurrent Fixed Bed ReactorJean Carlos Gonzalez HernandezNo ratings yet
- Properties of Oilfield WatersDocument7 pagesProperties of Oilfield WatersMaisam AbbasNo ratings yet
- 115 Rock Drill Oil ApplicationsDocument2 pages115 Rock Drill Oil ApplicationsRandi Riki AdtiaNo ratings yet
- OSU Biophysics Graduate HandbookDocument62 pagesOSU Biophysics Graduate HandbooksivaNo ratings yet
- Interactive Textbook 5 PDF Elelments 3 1Document5 pagesInteractive Textbook 5 PDF Elelments 3 1api-240094705No ratings yet
- T1 1 E Automotive 072Document15 pagesT1 1 E Automotive 072Marian OstrowskiNo ratings yet
- MSDS Colafax PmeDocument2 pagesMSDS Colafax Pmemndmatt100% (2)
- I Jet R Milk Adult RationDocument6 pagesI Jet R Milk Adult Rationmr cmfNo ratings yet
- Downstream Petroleum Sector and Fuel Quality StandardsDocument7 pagesDownstream Petroleum Sector and Fuel Quality Standardssanjayshah99No ratings yet
- ASTM A262, CorrosietestenDocument2 pagesASTM A262, CorrosietestenSales HBS Solutions100% (1)
- Dr. Fixit Piditop 333 Concrete Floor HardenerDocument3 pagesDr. Fixit Piditop 333 Concrete Floor Hardenerhunky11No ratings yet
- Datasheet For Steel Grades Structure Steel 16MnDocument2 pagesDatasheet For Steel Grades Structure Steel 16MnPond KampeeraNo ratings yet
- Tool Design TerminologyDocument5 pagesTool Design Terminologyanmol6237No ratings yet
- CBSE Sample Paper Class 7 Science Set 3Document5 pagesCBSE Sample Paper Class 7 Science Set 3Sumanta Pan100% (1)