You might also like
- Larvicidal Property of Sugar Apple (Annona Squamosa) and San Francisco (Codiaeum Variegatum) Against Yellow Fever Mosquito (Aedes Aegypti)Document4 pagesLarvicidal Property of Sugar Apple (Annona Squamosa) and San Francisco (Codiaeum Variegatum) Against Yellow Fever Mosquito (Aedes Aegypti)Euan VegaNo ratings yet
- Dar Et Al-2014-F Hepatica Desarrollo de Generaciones de Redias en Infeccion de P ColumellaDocument7 pagesDar Et Al-2014-F Hepatica Desarrollo de Generaciones de Redias en Infeccion de P Columellacba_ssNo ratings yet
- Effect of Red Palm Weevil Rhynchophorus FerrugineuDocument8 pagesEffect of Red Palm Weevil Rhynchophorus FerrugineuAli AlbaitiNo ratings yet
- Host Weed Species Range of Meloidogyne Ethiopica Whitehead (Tylenchida: Meloidogynidae) Found in BrazilDocument7 pagesHost Weed Species Range of Meloidogyne Ethiopica Whitehead (Tylenchida: Meloidogynidae) Found in BrazilWendy SerraNo ratings yet
- 10.biological Control With The Fungi PenicilliumDocument12 pages10.biological Control With The Fungi PenicilliumImpact JournalsNo ratings yet
- Insecticidal Effect of Dittrichia Viscosa Lyophilized Epicuticular MaterialDocument6 pagesInsecticidal Effect of Dittrichia Viscosa Lyophilized Epicuticular MaterialIlija MileticNo ratings yet
- Insecticidal Activity of A Destruxin-Containing Extract of Metarhizium Brunneum Against Ceratitis Capitata (Diptera: Tephritidae)Document11 pagesInsecticidal Activity of A Destruxin-Containing Extract of Metarhizium Brunneum Against Ceratitis Capitata (Diptera: Tephritidae)ahmredNo ratings yet
- Effect of Temperature and Water Activity On in Vitro Germination of Monilinia SPPDocument8 pagesEffect of Temperature and Water Activity On in Vitro Germination of Monilinia SPPOana PotocNo ratings yet
- Jee102 0144Document8 pagesJee102 0144Vicente MottaNo ratings yet
- Ee/nvu 064Document8 pagesEe/nvu 064Rosmery Naupari AlvarezNo ratings yet
- Grabner Et Al-2014-A Case Study of Parasite Fgigantica in PcolumellaDocument7 pagesGrabner Et Al-2014-A Case Study of Parasite Fgigantica in Pcolumellacba_ssNo ratings yet
- Nguyen Thi Kim Hue Et Al. 2020 Isolation and Identification of Herbivorous Ciliates From Contaminated Microalgal CulturesDocument11 pagesNguyen Thi Kim Hue Et Al. 2020 Isolation and Identification of Herbivorous Ciliates From Contaminated Microalgal CulturesHue NguyenNo ratings yet
- In Vitro Efficacy of Entomopathogenic Nematodes (EPNS) Against Economically Important Insect-Pests of CauliflowerDocument9 pagesIn Vitro Efficacy of Entomopathogenic Nematodes (EPNS) Against Economically Important Insect-Pests of CauliflowerMamta AgarwalNo ratings yet
- 7183 D20 ParasiticEnteritisAssociatedwiththeCamallanidserpinematrachemys2015ARAVDocument6 pages7183 D20 ParasiticEnteritisAssociatedwiththeCamallanidserpinematrachemys2015ARAV5dymas pandu pratamaNo ratings yet
- Ballesteros Et Al. Fern Gazette, 17 Full PaperDocument10 pagesBallesteros Et Al. Fern Gazette, 17 Full PaperDaniel BallesterosNo ratings yet
- Journal of Invertebrate Pathology: Óscar Dembilio, Enrique Quesada-Moraga, Cándido Santiago-Álvarez, Josep A. JacasDocument8 pagesJournal of Invertebrate Pathology: Óscar Dembilio, Enrique Quesada-Moraga, Cándido Santiago-Álvarez, Josep A. JacasEuclides DamiãoNo ratings yet
- 024 097 0428 PDFDocument8 pages024 097 0428 PDFDaniel AlzateNo ratings yet
- Biodiversity of Fungi On The Palm Eleiodoxa Conferta in Sirindhorn Peat Swamp Forest, Narathiwat, ThailandDocument14 pagesBiodiversity of Fungi On The Palm Eleiodoxa Conferta in Sirindhorn Peat Swamp Forest, Narathiwat, ThailanddhytaNo ratings yet
- BotryosphaeriaDocument10 pagesBotryosphaeriadavideNo ratings yet
- Environmental Persistence of Amphibian and Reptilian RanavirusesDocument8 pagesEnvironmental Persistence of Amphibian and Reptilian RanavirusesJamesComeyJustaBitchNo ratings yet
- Low-Dose Irradiation With Modified Atmosphere Packaging For Mango Against The Oriental Fruit Fly (Diptera: Tephritidae)Document6 pagesLow-Dose Irradiation With Modified Atmosphere Packaging For Mango Against The Oriental Fruit Fly (Diptera: Tephritidae)Haidar Ali AsshiddiqinNo ratings yet
- Frankliniella Occidentalis ManagementDocument7 pagesFrankliniella Occidentalis ManagementDani KurzelukNo ratings yet
- ZOJ 2024paratrichodorusallius-Complex RedDocument44 pagesZOJ 2024paratrichodorusallius-Complex RedccruzvasconcelosNo ratings yet
- Biological Control: V. San Andrés, I. Ayala, M.C. Abad, J. Primo, P. Castañera, P. MoyaDocument8 pagesBiological Control: V. San Andrés, I. Ayala, M.C. Abad, J. Primo, P. Castañera, P. MoyafadwaeNo ratings yet
- Animals 13 01764Document17 pagesAnimals 13 01764Ricardo HernándezNo ratings yet
- Ectoparasites of The Critically Endangered Insular Cavy, Cavia Intermedia (Rodentia: Caviidae), Southern BrazilDocument6 pagesEctoparasites of The Critically Endangered Insular Cavy, Cavia Intermedia (Rodentia: Caviidae), Southern Brazilf9cpdsfh6yNo ratings yet
- Heliyon: Shpend Shahini, Ajten B Erxolli, Frans KokojkaDocument7 pagesHeliyon: Shpend Shahini, Ajten B Erxolli, Frans KokojkaDevita AmeliaNo ratings yet
- Revista Mexicana de BiodiversidadDocument9 pagesRevista Mexicana de BiodiversidadPamela AlvarezNo ratings yet
- 14823-Article Text-47938-1-10-20191213Document10 pages14823-Article Text-47938-1-10-20191213Ghecrea GrigoreNo ratings yet
- 14823-Article Text-47938-1-10-20191213Document10 pages14823-Article Text-47938-1-10-20191213Ghecrea GrigoreNo ratings yet
- Susceptibility of Euschistus Heros and Dichelops FurcatusDocument8 pagesSusceptibility of Euschistus Heros and Dichelops Furcatusanapaula.mouraNo ratings yet
- Acta Tropica: Yersinia Pestis in Ectoparasites of Endemic and Domestic Animals in SouthwestDocument10 pagesActa Tropica: Yersinia Pestis in Ectoparasites of Endemic and Domestic Animals in SouthwestDiego TulcanNo ratings yet
- 2013 Assessment On The Diversity of paraDocument6 pages2013 Assessment On The Diversity of paraFAIRUZ KHALIDNo ratings yet
- Sociobiology: Research Article - WaspsDocument9 pagesSociobiology: Research Article - WaspsÍtalo SilvaNo ratings yet
- Eugenia Uniflora9Document6 pagesEugenia Uniflora9amensetNo ratings yet
- Revisión de Cambios - Artículo LinaDocument17 pagesRevisión de Cambios - Artículo LinaALEXNo ratings yet
- Gastrodiscoidiasis, A Plant-Borne Zoonotic Disease Caused by The Intestinal Amphistome Fluke Gastrodiscoides Hominis (Trematoda: Gastrodiscidae)Document7 pagesGastrodiscoidiasis, A Plant-Borne Zoonotic Disease Caused by The Intestinal Amphistome Fluke Gastrodiscoides Hominis (Trematoda: Gastrodiscidae)Hœ NęÿNo ratings yet
- Mortalidade de InsetosDocument2 pagesMortalidade de InsetosEloísaNo ratings yet
- Insects: Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in PakistanDocument12 pagesInsects: Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistanrida fatimaNo ratings yet
- Mycoendophytes As Pest Control Agents, Using Culture Filtrates, Against Green Apple Aphid (Hemiptera, Aphididae)Document11 pagesMycoendophytes As Pest Control Agents, Using Culture Filtrates, Against Green Apple Aphid (Hemiptera, Aphididae)Cedrus CedrusNo ratings yet
- Bioscientific Review (BSR) ::: HTTPS://SSC - Umt.Edu - Pk/Biosci/Home - AspxDocument11 pagesBioscientific Review (BSR) ::: HTTPS://SSC - Umt.Edu - Pk/Biosci/Home - AspxUMT JournalsNo ratings yet
- Khaldi Et Al. (2021) First Report and Infestation Levels of Zaprionus Indianus in AlgeriaDocument7 pagesKhaldi Et Al. (2021) First Report and Infestation Levels of Zaprionus Indianus in AlgeriaMourad KhaldiNo ratings yet
- Artículo - Evaluation of The Effectiveness of Dicyphus Errans As PredatorDocument7 pagesArtículo - Evaluation of The Effectiveness of Dicyphus Errans As PredatorAndrea Tafur AcostaNo ratings yet
- BICO 10.1007 s10526-015-9704-1Document11 pagesBICO 10.1007 s10526-015-9704-1Halisa IndrianiNo ratings yet
- Long-Term Studies On The Evolution of Resistance of Myzus Persicae (Hemiptera: Aphididae) To Insecticides in GreeceDocument16 pagesLong-Term Studies On The Evolution of Resistance of Myzus Persicae (Hemiptera: Aphididae) To Insecticides in Greeceammar47No ratings yet
- Torres Et Al. - 2006 - The Predatory Stinkbug Podisus Nigrispinus - Biology, Ecology and Augmentative Releases For Lepidopteran Larval Control in I Forests in Brazil.Document19 pagesTorres Et Al. - 2006 - The Predatory Stinkbug Podisus Nigrispinus - Biology, Ecology and Augmentative Releases For Lepidopteran Larval Control in I Forests in Brazil.Nilu Soofi Leon RubioNo ratings yet
- Harmful Algae: A B A A BDocument12 pagesHarmful Algae: A B A A BVictor A. Cervantes-UrietaNo ratings yet
- Navarro Et Al., 2019. ForesisDocument7 pagesNavarro Et Al., 2019. ForesismjnlNo ratings yet
- 84.2022 Coccinellidae Manabi BailonDocument7 pages84.2022 Coccinellidae Manabi BailonDorys ChirinosNo ratings yet
- Ectoparasitic Flies (Diptera: Streblidae) On Bats (Mammalia: Chiroptera) From A Private Natural Heritage Reserve in Southeastern BrazilDocument6 pagesEctoparasitic Flies (Diptera: Streblidae) On Bats (Mammalia: Chiroptera) From A Private Natural Heritage Reserve in Southeastern BrazilDaniel SantosNo ratings yet
- Fusarium Spp. in Loggerhead Sea Turtles (Caretta Caretta) : From Colonization To InfectionDocument8 pagesFusarium Spp. in Loggerhead Sea Turtles (Caretta Caretta) : From Colonization To InfectionPaz LibrosNo ratings yet
- Artigo Tricho 2Document8 pagesArtigo Tricho 2milenaaghata4No ratings yet
- Parasitic Interactions Between Nosema Spp. and Varroa Destructor in ApisDocument10 pagesParasitic Interactions Between Nosema Spp. and Varroa Destructor in ApisFrontiersNo ratings yet
- Basset & Lamarre 2019 ScienceDocument3 pagesBasset & Lamarre 2019 ScienceRutNo ratings yet
- Nota Técnica: ResumenDocument12 pagesNota Técnica: ResumenLuz Stella MurilloNo ratings yet
- Journal Homepage: - : IntroductionDocument9 pagesJournal Homepage: - : IntroductionIJAR JOURNALNo ratings yet
- 2019 Plant Disease FinalDocument10 pages2019 Plant Disease FinalKenny UriarteNo ratings yet
- Gyac 072Document12 pagesGyac 072I2O42OO54 IRFAN FAUZINo ratings yet
- Prevalence and Etiology of Dermatomycoses in Rajshahi, Bangladesh Farzana Ashrafi NeelaDocument4 pagesPrevalence and Etiology of Dermatomycoses in Rajshahi, Bangladesh Farzana Ashrafi Neelaabznaim420No ratings yet
- Nematodes and the Biological Control of Insect PestsFrom EverandNematodes and the Biological Control of Insect PestsRA BeddingRating: 1 out of 5 stars1/5 (1)
- Roses For Everyone Pitch Deck by SlidesgoDocument53 pagesRoses For Everyone Pitch Deck by Slidesgoatihrah kamarudinNo ratings yet
- Shamouti Orange IrrigationDocument6 pagesShamouti Orange IrrigationGissell Marisol Huamani ChavezNo ratings yet
- Short Communication Basal Media For in Vitro Germination of Red-Purple Dragon Fruit HylocereusDocument7 pagesShort Communication Basal Media For in Vitro Germination of Red-Purple Dragon Fruit Hylocereusjunior luis coronel maldonadoNo ratings yet
- Articulo 1. Persistencia de Plasmodiophora BrassicaeDocument27 pagesArticulo 1. Persistencia de Plasmodiophora Brassicaejunior luis coronel maldonadoNo ratings yet
- WPS Chapter 7Document34 pagesWPS Chapter 7richemengNo ratings yet
- Cylindicator Sensor Design GuideDocument2 pagesCylindicator Sensor Design GuideTavo VergaraNo ratings yet
- Chapter Seven: GS Ohlins Installation, Adjustment Tips & TricksDocument29 pagesChapter Seven: GS Ohlins Installation, Adjustment Tips & TricksWhattonNo ratings yet
- Manually Updating The Firmwares On A Dell PowerEdge R710 - Atualizar Firmwares DellDocument6 pagesManually Updating The Firmwares On A Dell PowerEdge R710 - Atualizar Firmwares Dellaltero spNo ratings yet
- ANFISDocument19 pagesANFISShimaa Barakat100% (1)
- Chloride LINEAR MK II - Service ManualDocument32 pagesChloride LINEAR MK II - Service Manualfabio.perazzoloNo ratings yet
- SMD TR60-1.0mm With 12mm Bar (Office Floor 3.64m Span) PDFDocument2 pagesSMD TR60-1.0mm With 12mm Bar (Office Floor 3.64m Span) PDFabhijitkolheNo ratings yet
- PRINCIPLES OF SURGERY (James R. Hupp Chapter 3 Notes) : 1. Develop A Surgical DiagnosisDocument5 pagesPRINCIPLES OF SURGERY (James R. Hupp Chapter 3 Notes) : 1. Develop A Surgical DiagnosisSonia LeeNo ratings yet
- The Sacred Number Forty-NineDocument12 pagesThe Sacred Number Forty-NinePatrick Mulcahy100% (6)
- V-Belt Sizing and Selection Guide: Standard V Belt Sizes FHP (Fractional Horsepower) V-Belts 3L, 4L, 5LDocument1 pageV-Belt Sizing and Selection Guide: Standard V Belt Sizes FHP (Fractional Horsepower) V-Belts 3L, 4L, 5LVijay ParmarNo ratings yet
- Jain 2018Document10 pagesJain 2018Pablo Ignacio Contreras EstradaNo ratings yet
- StructSteel Takeoff InstructionsDocument56 pagesStructSteel Takeoff InstructionsAmrish TyagiNo ratings yet
- EEMDocument17 pagesEEMSandaruwan සුජීවNo ratings yet
- Studyprotocol Open Access: Yue Yan, Yalin Zhan, Xian 'E Wang and Jianxia HouDocument7 pagesStudyprotocol Open Access: Yue Yan, Yalin Zhan, Xian 'E Wang and Jianxia HouGery KrismawanNo ratings yet
- M2-Exam 2Document13 pagesM2-Exam 2youngturtlevibesNo ratings yet
- C & Embedded QuestionsDocument109 pagesC & Embedded QuestionskalkikaliNo ratings yet
- Particle Tracing Module Users GuideDocument306 pagesParticle Tracing Module Users GuideAlfonso BarbozaNo ratings yet
- Univ of Michigan Flint Design Report 442Document74 pagesUniv of Michigan Flint Design Report 442Extra TerrestrialNo ratings yet
- Training - Manual - Diagnostic - and - Troublesh CPC 202GDocument186 pagesTraining - Manual - Diagnostic - and - Troublesh CPC 202GGA AnnerNo ratings yet
- 100 Irregular Plural Nouns ListDocument3 pages100 Irregular Plural Nouns Listjinesham007No ratings yet
- UACE BIO PAPER TWO SET 3 2023-JusanDocument3 pagesUACE BIO PAPER TWO SET 3 2023-JusanCampbell OGENRWOTNo ratings yet
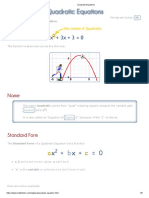
- Quadratic Equation - MATH IS FUNDocument8 pagesQuadratic Equation - MATH IS FUNChanchan LebumfacilNo ratings yet
- Bu3 Lecture1 Fundamentals of AcousticsDocument10 pagesBu3 Lecture1 Fundamentals of AcousticsPrincess HernandezNo ratings yet
- Irjet V4i10201 PDFDocument8 pagesIrjet V4i10201 PDFBesmir IsmailiNo ratings yet
- Heat Exchanger-Design and ConstructionDocument46 pagesHeat Exchanger-Design and ConstructionMohd Jamal Mohd Moktar100% (2)
- EECS4421Z: Introduction To Robotics Sample Exam QuestionsDocument7 pagesEECS4421Z: Introduction To Robotics Sample Exam QuestionsMooeez BellaamineNo ratings yet
- Hook Height With Drawwork SensorDocument3 pagesHook Height With Drawwork SensorJose PalumboNo ratings yet
- Drill Pipe Drill StringDocument11 pagesDrill Pipe Drill StringRodrigoBernardo50% (2)
- 2.1.2 Pseudocode PDFDocument13 pages2.1.2 Pseudocode PDFBlessed Endibye KanyembaNo ratings yet
- What Is A Philosophical Problem - HackerDocument12 pagesWhat Is A Philosophical Problem - Hackersnain1No ratings yet