You might also like
- The Enzyme Treatment of Cancer - John Beard 1911Document156 pagesThe Enzyme Treatment of Cancer - John Beard 1911Trombonino100% (18)
- Nephrotic SyndromeDocument13 pagesNephrotic SyndromeImanuel Far-FarNo ratings yet
- Out 28Document18 pagesOut 28EndraNo ratings yet
- 2008 - Antituberculosis Drug-Induced Hepatotoxicity Concise Up-To-Date ReviewDocument12 pages2008 - Antituberculosis Drug-Induced Hepatotoxicity Concise Up-To-Date ReviewCarlos LuqueNo ratings yet
- Drug Treatment in TuberculosisDocument8 pagesDrug Treatment in TuberculosisCaren ChanNo ratings yet
- Jurnal MDR TBDocument7 pagesJurnal MDR TBIndra HebatNo ratings yet
- Jurnal BioselDocument15 pagesJurnal BioselRegita cahyani priscalisyiNo ratings yet
- Recent Advances in The Development of Anti-Tuberculosis Drugs Acting On Multidrug-Resistant Strains: A ReviewDocument18 pagesRecent Advances in The Development of Anti-Tuberculosis Drugs Acting On Multidrug-Resistant Strains: A Reviewmalik003No ratings yet
- Potent TgCDPK1 Inhibitors with Anti-Toxoplasma ActivityDocument25 pagesPotent TgCDPK1 Inhibitors with Anti-Toxoplasma ActivityVivek Ananth R PNo ratings yet
- Fasting Enhances Cancer TherapyDocument9 pagesFasting Enhances Cancer TherapyNurul Dwi Utami100% (1)
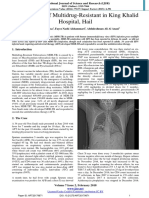
- MDR-TB HIV Coinfection Case Report King Khalid HospitalDocument2 pagesMDR-TB HIV Coinfection Case Report King Khalid HospitalryoNo ratings yet
- Anti Cancer DrugsDocument56 pagesAnti Cancer DrugsTimothy David50% (2)
- Ijms 23 08337 v2Document15 pagesIjms 23 08337 v2응우엔티 트랑No ratings yet
- antifungal pdfDocument34 pagesantifungal pdfUmashankar SharmaNo ratings yet
- Unit - 3Document39 pagesUnit - 3ujjwalNo ratings yet
- Gostner Et AlDocument11 pagesGostner Et Algiulia.santinNo ratings yet
- Suliman 2018Document12 pagesSuliman 2018juheb ansariNo ratings yet
- Mycoplasma Pneumoniae ThesisDocument7 pagesMycoplasma Pneumoniae Thesistoniawallacelittlerock100% (2)
- MDR TBDocument8 pagesMDR TBMahira Bayu AdiftaNo ratings yet
- FilariasisDocument13 pagesFilariasisJohan Stiven Mahecha CobosNo ratings yet
- Typhoid FeverDocument6 pagesTyphoid FeverReeshabhdev GauttamNo ratings yet
- Mitochondrial Unfolded Protein ResponseDocument10 pagesMitochondrial Unfolded Protein Responseنوره نورNo ratings yet
- Yuandani 2013Document10 pagesYuandani 2013Khrisna Whaty SilalahiNo ratings yet
- Itpa Polymorphisms and The Incidence of ToxicitiesDocument6 pagesItpa Polymorphisms and The Incidence of ToxicitiesChen JesseNo ratings yet
- Audiologic Monitoring MDR TBDocument7 pagesAudiologic Monitoring MDR TBLussie_varetaNo ratings yet
- GMN MembranosaDocument23 pagesGMN MembranosaGabriela VaraNo ratings yet
- Immunosuppressive Drug TherapyDocument25 pagesImmunosuppressive Drug Therapyapi-3741769100% (2)
- Winnicki MPADocument12 pagesWinnicki MPAMonikaNo ratings yet
- Adverse Event Profile of Pyrimethamine-Based Therapy in Toxoplasmosis: A Systematic ReviewDocument22 pagesAdverse Event Profile of Pyrimethamine-Based Therapy in Toxoplasmosis: A Systematic Reviewirawati hidayahNo ratings yet
- Review On Pulse Therapy A Novel Approach in The Treatment of Pemphigus Vulgaris PDFDocument4 pagesReview On Pulse Therapy A Novel Approach in The Treatment of Pemphigus Vulgaris PDFtaniaNo ratings yet
- Bereda 2022Document10 pagesBereda 2022mdvalentNo ratings yet
- Cancers 15 03408 v2Document14 pagesCancers 15 03408 v2Jesus Barraza SánchezNo ratings yet
- Zhou2008 Vis DujDocument47 pagesZhou2008 Vis DujAnAn BanhGaoNo ratings yet
- Articulo OooDocument7 pagesArticulo OooPaola M. TrianaNo ratings yet
- New JaneDocument34 pagesNew JaneniputusellyowNo ratings yet
- Apigenin Parsley Melanoma PD-1 PD-L1Document15 pagesApigenin Parsley Melanoma PD-1 PD-L1valgurNo ratings yet
- Plasmodium Falciparum Dihydroorotate Dehydrogenase: A Drug Target Against MalariaDocument24 pagesPlasmodium Falciparum Dihydroorotate Dehydrogenase: A Drug Target Against MalariaDelvina RahmadaniNo ratings yet
- In-Silico Drug Design Approach For Identification of Antibacterial Drug Leads For Treatment of Gastric LymphomaDocument7 pagesIn-Silico Drug Design Approach For Identification of Antibacterial Drug Leads For Treatment of Gastric LymphomaWARSE JournalsNo ratings yet
- 6.Challenges in Chagas disease drug developmentDocument14 pages6.Challenges in Chagas disease drug developmentyerli bahosNo ratings yet
- Purine Salvage Pathway in Mycobacterium Tuberculosis: R.G. Ducati, A. Breda, L.A. Basso and D.S. SantosDocument18 pagesPurine Salvage Pathway in Mycobacterium Tuberculosis: R.G. Ducati, A. Breda, L.A. Basso and D.S. SantosDeepabali GhosalNo ratings yet
- Lister Strain Vaccinia Virus, A Potential Therapeutic Vector Targeting Hypoxic TumoursDocument15 pagesLister Strain Vaccinia Virus, A Potential Therapeutic Vector Targeting Hypoxic TumourssusasureshNo ratings yet
- Once-Daily Plazomicin For Complicated Urinary Tract InfectionsDocument12 pagesOnce-Daily Plazomicin For Complicated Urinary Tract InfectionsharvardboyNo ratings yet
- Role of Hypoxia in Cancer Therapy by Regulating The Tumor MicroenvironmentDocument15 pagesRole of Hypoxia in Cancer Therapy by Regulating The Tumor MicroenvironmentShuang LiNo ratings yet
- مقالههههDocument34 pagesمقالههههyosof esfandiariNo ratings yet
- Grupo 5 Metabolismo Del Triptofano en PsiconeuroinmunologiaDocument11 pagesGrupo 5 Metabolismo Del Triptofano en PsiconeuroinmunologiaDaniel J Alejos CNo ratings yet
- Journal Pre-Proof: European Journal of Medicinal ChemistryDocument28 pagesJournal Pre-Proof: European Journal of Medicinal ChemistryColo VoltaNo ratings yet
- Fonc 09 00396Document8 pagesFonc 09 00396asdffdsaNo ratings yet
- Multi-Drug-resistant Tuberculosis - Wikipedia, The Free EncyclopediaDocument8 pagesMulti-Drug-resistant Tuberculosis - Wikipedia, The Free Encyclopedianayan555No ratings yet
- Emerging Treatments For Crohn's Disease: Cells, Surgery, and Novel TherapeuticsDocument10 pagesEmerging Treatments For Crohn's Disease: Cells, Surgery, and Novel Therapeuticsellya theresiaNo ratings yet
- Introduction to Antimicrobial Chemotherapy: Mechanisms and SpectrumDocument31 pagesIntroduction to Antimicrobial Chemotherapy: Mechanisms and SpectrumPrasad SangishettyNo ratings yet
- Medical Care: "Microbiology, Epidemiology, Clinical Manifestations, and Diagnosis of Leptospirosis"Document5 pagesMedical Care: "Microbiology, Epidemiology, Clinical Manifestations, and Diagnosis of Leptospirosis"Lyra LorcaNo ratings yet
- Jurnal 1 Kelompok 9Document17 pagesJurnal 1 Kelompok 9hy brezzleNo ratings yet
- MDR TB MehakDocument6 pagesMDR TB MehakKeham LeogNo ratings yet
- 24 OctDocument38 pages24 OctVenkatesh JaiswalNo ratings yet
- Defenisi PDFDocument2 pagesDefenisi PDFNyoman SuryaNo ratings yet
- 08Document11 pages08gvignesh2629No ratings yet
- Researcharticle Open AccessDocument13 pagesResearcharticle Open AccessManuela Martín-Bejarano GarcíaNo ratings yet
- Recent Advances in Antiviral Therapy: Derek KinchingtonDocument7 pagesRecent Advances in Antiviral Therapy: Derek KinchingtonYogesh RathiNo ratings yet
- Tumor Immune Microenvironment in Cancer Progression and Cancer TherapyFrom EverandTumor Immune Microenvironment in Cancer Progression and Cancer TherapyPawel KalinskiNo ratings yet
- The Basics of Cancer ImmunotherapyFrom EverandThe Basics of Cancer ImmunotherapyHaidong DongNo ratings yet
- Infections in Cancer Chemotherapy: A Symposium Held at the Institute Jules Bordet, Brussels, BelgiumFrom EverandInfections in Cancer Chemotherapy: A Symposium Held at the Institute Jules Bordet, Brussels, BelgiumNo ratings yet
- 2017 Article 1797 PDFDocument5 pages2017 Article 1797 PDFluanadumitruNo ratings yet
- Thomas McPherson Brown MD Treatment of Rheumatoid DiseaseDocument29 pagesThomas McPherson Brown MD Treatment of Rheumatoid DiseaseLidia Lidia100% (1)
- Prsentation LA 1 Basics and External Laser Therapy PDFDocument197 pagesPrsentation LA 1 Basics and External Laser Therapy PDFluanadumitruNo ratings yet
- EMERGE Plague Recommendations PDFDocument6 pagesEMERGE Plague Recommendations PDFluanadumitruNo ratings yet
- J. Nutr.-2004-Wu-489-92Document4 pagesJ. Nutr.-2004-Wu-489-92Luis Marcas VilaNo ratings yet
- Nihms716157 PDFDocument16 pagesNihms716157 PDFFaisal RuhiatnaNo ratings yet
- Nihms716157 PDFDocument16 pagesNihms716157 PDFFaisal RuhiatnaNo ratings yet
- Novel Functions of Vitamin b6 PDFDocument6 pagesNovel Functions of Vitamin b6 PDFluanadumitruNo ratings yet
- Cyp450 Nutrient Inhibitors & Inducers: The Insider's Guide To TheDocument21 pagesCyp450 Nutrient Inhibitors & Inducers: The Insider's Guide To TheluanadumitruNo ratings yet
- History of Selenium Research 15 Jan 2017-4 PDFDocument103 pagesHistory of Selenium Research 15 Jan 2017-4 PDFluanadumitruNo ratings yet
- Am J Clin Nutr-2007-Maintz-1185-96 PDFDocument12 pagesAm J Clin Nutr-2007-Maintz-1185-96 PDFluanadumitruNo ratings yet
- Resume of Intravenous Procaine Therapy PDFDocument6 pagesResume of Intravenous Procaine Therapy PDFluanadumitruNo ratings yet
- Photobiomodulation Compressed PDFDocument27 pagesPhotobiomodulation Compressed PDFluanadumitruNo ratings yet
- History of Selenium Research 15 Jan 2017-4 PDFDocument103 pagesHistory of Selenium Research 15 Jan 2017-4 PDFluanadumitruNo ratings yet
- Newborn Pics SkinDocument56 pagesNewborn Pics SkinluanadumitruNo ratings yet
- Resume of Intravenous Procaine Therapy PDFDocument6 pagesResume of Intravenous Procaine Therapy PDFluanadumitruNo ratings yet
- Novel Functions of Vitamin b6 PDFDocument6 pagesNovel Functions of Vitamin b6 PDFluanadumitruNo ratings yet
- Protein Contents of Plant FoodsDocument3 pagesProtein Contents of Plant FoodsluanadumitruNo ratings yet
- Herbs Leaky GutDocument7 pagesHerbs Leaky GutMaria Bulacu100% (1)
- BME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDocument67 pagesBME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDewi Cahya FitriNo ratings yet
- 09-08-15 SR - Iit-Iz-Co-Spark Jee Adv Rpta-2 (2013 p2) Q'paperDocument33 pages09-08-15 SR - Iit-Iz-Co-Spark Jee Adv Rpta-2 (2013 p2) Q'paperKumar Prasad100% (1)
- FlyerDocument2 pagesFlyerCamilo Galindo CaldasNo ratings yet
- Report of Micro Project (2) - 1Document8 pagesReport of Micro Project (2) - 1ythNo ratings yet
- DAE SSPS 2004 PresentationDocument32 pagesDAE SSPS 2004 PresentationRavi Kumar ChNo ratings yet
- White1971 - Vapor Dominated Hydrotermal Systems Compared With Hot-Water SystemDocument23 pagesWhite1971 - Vapor Dominated Hydrotermal Systems Compared With Hot-Water SystemNino PumaNo ratings yet
- CBM 2016 1Document14 pagesCBM 2016 1SHEIKH MUHAMMAD FAHADNo ratings yet
- Chemistry of Transition MetalsDocument56 pagesChemistry of Transition MetalsHamad Mohamad100% (1)
- Kilfrost Winter Best Practice Guide - Edition 3Document44 pagesKilfrost Winter Best Practice Guide - Edition 3DimitriosMonogios100% (1)
- Testing of Building Stones - Rock Tests On Lab & FieldDocument3 pagesTesting of Building Stones - Rock Tests On Lab & FieldআকাশআহসানNo ratings yet
- Science Revision Sheet - Term 1-Year 4: NameDocument18 pagesScience Revision Sheet - Term 1-Year 4: NameSaid HusseinNo ratings yet
- Gear Beam and Wear StrengthDocument46 pagesGear Beam and Wear StrengthNisarg MistryNo ratings yet
- Assessment Modules 5 6Document3 pagesAssessment Modules 5 6Obeña Grecia Joy T.No ratings yet
- Temperature and Heat 3Document18 pagesTemperature and Heat 3Samud MorrisNo ratings yet
- The Diffusion of Gases Through Fused QuartzDocument6 pagesThe Diffusion of Gases Through Fused QuartzElenaNo ratings yet
- Improving Separation of Basic Compounds by Reversed-Phase HPLC Using Chaotropic AgentsDocument7 pagesImproving Separation of Basic Compounds by Reversed-Phase HPLC Using Chaotropic AgentsKriti TyagiNo ratings yet
- MSDSDocument13 pagesMSDSSehry SyedNo ratings yet
- Safety Data Sheet Highlights Hazards of Hostapon SCI 85 GDocument10 pagesSafety Data Sheet Highlights Hazards of Hostapon SCI 85 GFirsta Sarascia VindyNo ratings yet
- Board Exam Review - Set 3BDocument17 pagesBoard Exam Review - Set 3BJey-r VenturaNo ratings yet
- Water Reclamation and Reuse: Huijuan Sun, Huixin Zhang, Rui Xu, Xin Zou and Yang LiuDocument21 pagesWater Reclamation and Reuse: Huijuan Sun, Huixin Zhang, Rui Xu, Xin Zou and Yang LiujheufguwrfhNo ratings yet
- Air Entraining CementDocument14 pagesAir Entraining CementBashairu WaseemNo ratings yet
- Petrofecta: Automated Trapped Fluid and Elemental Composition AnalysisDocument1 pagePetrofecta: Automated Trapped Fluid and Elemental Composition AnalysisRifan MahulauwNo ratings yet
- Freon 134a Si Thermodynamic PropertiesDocument52 pagesFreon 134a Si Thermodynamic PropertiesAyu LestariNo ratings yet
- Adhesion of Cells To Polystyrene SurfaceDocument7 pagesAdhesion of Cells To Polystyrene Surfacef20212314No ratings yet
- 8 Engine Terms Every Marine Engineer Should Know - Part 2Document9 pages8 Engine Terms Every Marine Engineer Should Know - Part 2tomNo ratings yet
- Determinación de Acrilamida en Dieferentes MaterialesDocument88 pagesDeterminación de Acrilamida en Dieferentes MaterialesMarianella ZegarraNo ratings yet
- ST Epan: Product BulletinDocument2 pagesST Epan: Product BulletinJulio Ramos GonzalezNo ratings yet
- Light Rigidizable Inflatable Wings For UAVs Resin and Manufacturing DevelopmentDocument16 pagesLight Rigidizable Inflatable Wings For UAVs Resin and Manufacturing DevelopmentMohd AnuarNo ratings yet
- GenBio 1 - 2nd Quarter ReviewerDocument8 pagesGenBio 1 - 2nd Quarter Reviewerjoshua tejadaNo ratings yet
- Particulate Filled Polypropylene: Structure and PropertiesDocument70 pagesParticulate Filled Polypropylene: Structure and PropertiesMinh NguyễnNo ratings yet