You might also like
- Immunoinformatic Analysis of Sars-Cov-2 Nucleocapsid Protein and Identi Fication of Covid-19 Vaccine TargetsDocument10 pagesImmunoinformatic Analysis of Sars-Cov-2 Nucleocapsid Protein and Identi Fication of Covid-19 Vaccine TargetsThor RagnarokNo ratings yet
- Flavonoid Covid 19Document13 pagesFlavonoid Covid 19nadia firaNo ratings yet
- In SilicoDocument9 pagesIn SilicoShafa ShaviraNo ratings yet
- Bioactive Molecules Derived From Snake Venoms With Therapeutic Potential For The Treatment of Thrombo-Cardiovascular Disorders Associated With COVID-19Document43 pagesBioactive Molecules Derived From Snake Venoms With Therapeutic Potential For The Treatment of Thrombo-Cardiovascular Disorders Associated With COVID-19emmanuel AndemNo ratings yet
- Investigación - CovidDocument12 pagesInvestigación - CovidMauricio FemeniaNo ratings yet
- SARS Cov2 and Ubiquitin PathwayDocument13 pagesSARS Cov2 and Ubiquitin PathwayHooraina Hassan SiddiquiNo ratings yet
- Plasmin Cascade Mediates Thrombotic Events in Sars-Cov-2 Infection Via Complement and Platelet-Activating SystemsDocument8 pagesPlasmin Cascade Mediates Thrombotic Events in Sars-Cov-2 Infection Via Complement and Platelet-Activating SystemsGF MANo ratings yet
- Journal Pre-Proof: Uhlberger, Jason D. GraciDocument43 pagesJournal Pre-Proof: Uhlberger, Jason D. GraciMohammed Shuaib AhmedNo ratings yet
- Sarscov2 Promote InflammationDocument13 pagesSarscov2 Promote InflammationAgnese ValentiniNo ratings yet
- Deciphering COVID-19 A ReviewDocument12 pagesDeciphering COVID-19 A ReviewInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- 10 1016@j Ijbiomac 2020 06 228Document8 pages10 1016@j Ijbiomac 2020 06 228Wahyu dickyNo ratings yet
- Covid PGXDocument13 pagesCovid PGXDyahsekarayupudak wangiNo ratings yet
- 2020 Distinct Infection Process of SARS CoV 2 in Human Bronchial Epithelial Cell LinesDocument9 pages2020 Distinct Infection Process of SARS CoV 2 in Human Bronchial Epithelial Cell LinesMartin JYNo ratings yet
- Cei 202 162Document31 pagesCei 202 162chaterinaNo ratings yet
- Bioinformatic Prediction of Potential T Cell Epitopes For SARS-Cov-2Document7 pagesBioinformatic Prediction of Potential T Cell Epitopes For SARS-Cov-2kjhgfghjkNo ratings yet
- Sars Cov2 Antigen Presentation Process by Immune Cells: Immunology Assignment - 2Document17 pagesSars Cov2 Antigen Presentation Process by Immune Cells: Immunology Assignment - 2azeema fatimaNo ratings yet
- Khalsa College, Amritsar: A Seminar On: Role of NK Cells in Sars-Cov-2Document20 pagesKhalsa College, Amritsar: A Seminar On: Role of NK Cells in Sars-Cov-2Arundhati VashishthNo ratings yet
- S1 Spike Protein Causes PASC, Cytokine Storm, & Long COVIDDocument115 pagesS1 Spike Protein Causes PASC, Cytokine Storm, & Long COVIDAnthony DocKek PenaNo ratings yet
- Melatonina Na COVIDDocument17 pagesMelatonina Na COVIDDiego SantanaNo ratings yet
- Overview JournalDocument8 pagesOverview JournalDea Inthay WulanNo ratings yet
- 5400-Article Text-31589-2-10-20210820Document12 pages5400-Article Text-31589-2-10-20210820Voltaire Méndez RodríguezNo ratings yet
- Sengupta Et Al., 2022 - COVID-19 & Vitamin CDocument8 pagesSengupta Et Al., 2022 - COVID-19 & Vitamin CDr.Pallav SenguptaNo ratings yet
- Caractere VirologiaueDocument29 pagesCaractere Virologiauehind el hamriNo ratings yet
- Sars-Cov-2 Evolution and Vaccines: Cause For Concern?: CommentDocument3 pagesSars-Cov-2 Evolution and Vaccines: Cause For Concern?: Commenthymen busterNo ratings yet
- Nac Covid Inflamación Daño Tisular 2020 NasiDocument4 pagesNac Covid Inflamación Daño Tisular 2020 NasiLuis C Ribon VNo ratings yet
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroMohamed Fawzy mahrousNo ratings yet
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in Vitromuun yayo100% (1)
- Cell Deaths - Involvement in The Pathogenesis and InterventionDocument21 pagesCell Deaths - Involvement in The Pathogenesis and InterventionSHREYA SENTHIL KUMAR IMS21286No ratings yet
- Measuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Document12 pagesMeasuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Chris KontomarisNo ratings yet
- Mahdian2020 Article DrugRepurposingUsingComputatioDocument9 pagesMahdian2020 Article DrugRepurposingUsingComputatioAndrew L GreearNo ratings yet
- Artículo PDFDocument11 pagesArtículo PDFKarlaMariaMartinezOrdoñezNo ratings yet
- 1 s2.0 S0753332220305308 MainDocument4 pages1 s2.0 S0753332220305308 Mainvictor rodriguezNo ratings yet
- Antioxidants: Glutathione Supplementation As An Adjunctive Therapy in COVID-19Document22 pagesAntioxidants: Glutathione Supplementation As An Adjunctive Therapy in COVID-19Felicia JesslynNo ratings yet
- Paper Jbuon Nov 21 (26-5-1719)Document4 pagesPaper Jbuon Nov 21 (26-5-1719)dimitriosroukasNo ratings yet
- Immune Response and Molecular Mechanisms of Cardiovascular Adverse Effects of Spike Proteins From SARS-CoV-2 and mRNA VaccinesDocument28 pagesImmune Response and Molecular Mechanisms of Cardiovascular Adverse Effects of Spike Proteins From SARS-CoV-2 and mRNA Vaccinesemmanuel AndemNo ratings yet
- Journal Pre-Proof: Cell Reports MedicineDocument39 pagesJournal Pre-Proof: Cell Reports MedicineErick RomeroNo ratings yet
- Covid 02 00104Document11 pagesCovid 02 00104Md. Rakibul IslamNo ratings yet
- Molecules 25 03822Document28 pagesMolecules 25 03822Ahmad KadafiNo ratings yet
- PacesJ2020 379 COVID 19andimmunesystem Rev PDFDocument11 pagesPacesJ2020 379 COVID 19andimmunesystem Rev PDFTaif RashedNo ratings yet
- COVID-19, Immune System Response, Hyperinflammation and Repurposing Antirheumatic DrugsDocument13 pagesCOVID-19, Immune System Response, Hyperinflammation and Repurposing Antirheumatic DrugsMarcelitaTaliaDuwiriNo ratings yet
- Sars-Cov-2 Seroconversion in Humans: A Detailed Protocol For A Serological Assay, Antigen Production, and Test SetupDocument15 pagesSars-Cov-2 Seroconversion in Humans: A Detailed Protocol For A Serological Assay, Antigen Production, and Test SetupAKNTAI002No ratings yet
- In Vitro and Animal Models For Sars-Cov-2 Research: ForumDocument5 pagesIn Vitro and Animal Models For Sars-Cov-2 Research: ForumJack SupramaniamNo ratings yet
- Journal Pre-Proof: Microbes and InfectionDocument42 pagesJournal Pre-Proof: Microbes and InfectionPaoloMafaldoNo ratings yet
- 1 s2.0 S075333222200261X MainDocument9 pages1 s2.0 S075333222200261X MainمولديياNo ratings yet
- PIIS0168827820336758Document17 pagesPIIS0168827820336758Yusup AhmadiNo ratings yet
- Bioorganic & Medicinal Chemistry Letters: Sven Ullrich, Christoph Nitsche TDocument8 pagesBioorganic & Medicinal Chemistry Letters: Sven Ullrich, Christoph Nitsche TMarcos SilvaNo ratings yet
- SARS-CoV-2 X ERDocument6 pagesSARS-CoV-2 X ERJulia MouraNo ratings yet
- Erythro VLPsDocument19 pagesErythro VLPsLucas HeNo ratings yet
- CimbDocument13 pagesCimbDr. Mohd AriffuddinNo ratings yet
- Proteomics Clinical Apps - 2023 - Brogna - Detection of Recombinant Spike Protein in The Blood of Individuals VaccinatedDocument7 pagesProteomics Clinical Apps - 2023 - Brogna - Detection of Recombinant Spike Protein in The Blood of Individuals Vaccinatedgoyapatez2No ratings yet
- s41423 021 00813 6Document12 pagess41423 021 00813 6Josué Cristhian Del Valle HornaNo ratings yet
- Processes: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensDocument11 pagesProcesses: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensHan VoNo ratings yet
- COVID 19 DrugsDocument9 pagesCOVID 19 DrugsTausif AlamNo ratings yet
- Ortegaa Et Al, 2020Document8 pagesOrtegaa Et Al, 2020Kassila SantosNo ratings yet
- Growth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationDocument28 pagesGrowth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationBf LiawNo ratings yet
- Banerjee Et Al 2020Document5 pagesBanerjee Et Al 2020Alex RomeroNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateRocio GmarNo ratings yet
- Respiratory MedicineDocument8 pagesRespiratory MedicineSky AngelNo ratings yet
- Kimia Medisinal: Oleh Kelompok 8Document16 pagesKimia Medisinal: Oleh Kelompok 8Hasryanto YantoNo ratings yet
- Medical Hypotheses: SciencedirectDocument8 pagesMedical Hypotheses: SciencedirectautomationenggNo ratings yet
- 1 s2.0 S1876034120304445 MainDocument5 pages1 s2.0 S1876034120304445 MainautomationenggNo ratings yet
- 1 s2.0 S0377123720300988 MainDocument6 pages1 s2.0 S0377123720300988 MainautomationenggNo ratings yet
- Articles: BackgroundDocument10 pagesArticles: BackgroundautomationenggNo ratings yet
- BBA - Molecular Basis of Disease: ReviewDocument12 pagesBBA - Molecular Basis of Disease: ReviewautomationenggNo ratings yet
- Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2Document10 pagesStructural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2automationenggNo ratings yet
- Sars-Cov-2: Camazotz 'S Curse: SciencedirectDocument6 pagesSars-Cov-2: Camazotz 'S Curse: SciencedirectautomationenggNo ratings yet
- European Journal of Medicinal ChemistryDocument12 pagesEuropean Journal of Medicinal ChemistryautomationenggNo ratings yet
- Science of The Total EnvironmentDocument5 pagesScience of The Total EnvironmentautomationenggNo ratings yet
- 1 s2.0 S0022510X20304226 Main PDFDocument18 pages1 s2.0 S0022510X20304226 Main PDFautomationenggNo ratings yet
- Gene Reports: SciencedirectDocument7 pagesGene Reports: SciencedirectautomationenggNo ratings yet
- European Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilDocument11 pagesEuropean Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilautomationenggNo ratings yet
- International Journal of Antimicrobial AgentsDocument9 pagesInternational Journal of Antimicrobial AgentsautomationenggNo ratings yet
- MSC Project Presentation 29 MarchDocument36 pagesMSC Project Presentation 29 MarchdreamroseNo ratings yet
- EPIDEMIOLOGY OF NON COMMUNICABLE DISEASES FinalDocument30 pagesEPIDEMIOLOGY OF NON COMMUNICABLE DISEASES FinalShashah RauNo ratings yet
- 2 4-Of My SiblingsDocument20 pages2 4-Of My SiblingsAbdul AzizNo ratings yet
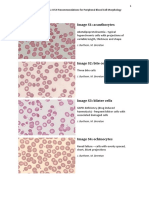
- ICSH Recommendations For Peripheral Blood Cell Morphology Standardization and GradingDocument9 pagesICSH Recommendations For Peripheral Blood Cell Morphology Standardization and GradingWilmer C. Geri Paredes100% (1)
- Puberty Suppression Medicine or MalpracticeDocument35 pagesPuberty Suppression Medicine or Malpracticebashomatsuo50% (2)
- Revision - Test - Topic - 7 - To - 17 MsDocument33 pagesRevision - Test - Topic - 7 - To - 17 MsZaigham ZiaNo ratings yet
- Dear Editor by Prof. Dr. Pir NasiruddaulaDocument147 pagesDear Editor by Prof. Dr. Pir Nasiruddaulabigleo4meNo ratings yet
- Experiment 11 - Digestion of FoodstuffsDocument7 pagesExperiment 11 - Digestion of FoodstuffsMark Ryan Tripole100% (1)
- Chromosome Structure and FunctionDocument76 pagesChromosome Structure and Functionstevensb055No ratings yet
- Renr Week 7th 2017 Questions SheetDocument21 pagesRenr Week 7th 2017 Questions SheetSasha UterNo ratings yet
- TraumaDocument10 pagesTraumadrhazimsalih100% (1)
- Insulin Pump Therapy Vs InjectionsDocument6 pagesInsulin Pump Therapy Vs InjectionsHakan KüçükerNo ratings yet
- E 3 FBDocument21 pagesE 3 FBvidya.damodaran3594No ratings yet
- Super Moringa EbookDocument43 pagesSuper Moringa Ebookanteparts100% (1)
- The Use of Hyaluronidase in Cosmetic DermatologyDocument7 pagesThe Use of Hyaluronidase in Cosmetic DermatologyDr. Hilder HernandezNo ratings yet
- TALENT EXsampleX50Document14 pagesTALENT EXsampleX50Kundan KumarNo ratings yet
- Measures of Diagnostic Accuracy: Basic Definitions: Ana-Maria ŠimundićDocument9 pagesMeasures of Diagnostic Accuracy: Basic Definitions: Ana-Maria ŠimundićgangjothelancerNo ratings yet
- SycosisDocument9 pagesSycosisVirag PatilNo ratings yet
- Disaster Nursing CompetenciesDocument84 pagesDisaster Nursing Competenciesdewisimoo100% (1)
- Christopher Columbus EssayDocument6 pagesChristopher Columbus Essayjvscmacaf100% (2)
- Chapter 3 Lesson 3 ProtozoaDocument17 pagesChapter 3 Lesson 3 ProtozoaKreizel FajaNo ratings yet
- Referat Gangguan Psikotik AkutDocument8 pagesReferat Gangguan Psikotik AkutYoga Oktavian NugrahaNo ratings yet
- Appendicitis Pregnancy ElnasharDocument47 pagesAppendicitis Pregnancy ElnasharErwynsonsaut SimanjuntakNo ratings yet
- Alexis - Protector-Retractor PlagaDocument8 pagesAlexis - Protector-Retractor PlagaMarius NenciuNo ratings yet
- Sarah S. Taupan, RN, MN, DPADocument33 pagesSarah S. Taupan, RN, MN, DPAKatri ArasaNo ratings yet
- Health Problems Common in ToddlersDocument4 pagesHealth Problems Common in ToddlersKim_b_Serac_6246No ratings yet
- Expt. 6 Biochemical Test of UrineDocument13 pagesExpt. 6 Biochemical Test of UrineMary Ella Mae PilaNo ratings yet
- Blepharoplasty ComplicationsDocument12 pagesBlepharoplasty ComplicationsMaría Alejandra Rojas MontenegroNo ratings yet
- Thesis On FundraisingDocument5 pagesThesis On Fundraisingk1lod0zolog3100% (2)
- Vinmec Services Marketing ReportDocument32 pagesVinmec Services Marketing ReportNgô Hồng Giang 2M-20ACN100% (1)