You might also like
- Potential Antiviral Agents for Coronaviruses: Compounds, Herbal Products, and Drug TargetsFrom EverandPotential Antiviral Agents for Coronaviruses: Compounds, Herbal Products, and Drug TargetsNo ratings yet
- Alternatives To Vancomycin For The Treatment of Methicillin-Resistant Staphylococcus Aureus InfectionsDocument7 pagesAlternatives To Vancomycin For The Treatment of Methicillin-Resistant Staphylococcus Aureus InfectionsInna MaynizaNo ratings yet
- Mscano, Journal Editor, Jidc-012-0978Document7 pagesMscano, Journal Editor, Jidc-012-0978imyours9601No ratings yet
- Daptomycin For The Treatment of Bacteraemia Du - 2014 - International Journal ofDocument9 pagesDaptomycin For The Treatment of Bacteraemia Du - 2014 - International Journal ofPatricio Suazo SotoNo ratings yet
- Buletin Farmasi 02/2013Document19 pagesBuletin Farmasi 02/2013afiq83No ratings yet
- البحث الاولDocument8 pagesالبحث الاولMarwa MostafaNo ratings yet
- Vancomycin-Resistant Enterococcal Infections: Epidemiology, Clinical Manifestations, and Optimal ManagementDocument14 pagesVancomycin-Resistant Enterococcal Infections: Epidemiology, Clinical Manifestations, and Optimal ManagementFernando Martínez HiguerasNo ratings yet
- Tristan O'Driscoll1 Christopher W Crank2Document14 pagesTristan O'Driscoll1 Christopher W Crank2HQ HQNo ratings yet
- Enterococcus 2Document8 pagesEnterococcus 2Sol BorturoNo ratings yet
- Estes-Derendorf2010 Article ComparisonOfThePharmacokineticDocument11 pagesEstes-Derendorf2010 Article ComparisonOfThePharmacokineticAyushi KapoorNo ratings yet
- Comparison of In-Vitro Efficacy of Vanc, LNZ and Dapto Angainst MRSA.Document6 pagesComparison of In-Vitro Efficacy of Vanc, LNZ and Dapto Angainst MRSA.Nawwal NaeemNo ratings yet
- European Journal of Medicinal ChemistryDocument14 pagesEuropean Journal of Medicinal ChemistryWalid Ebid ElgammalNo ratings yet
- Bio Sem Final Draft 1Document15 pagesBio Sem Final Draft 1api-298963157No ratings yet
- Fosfomycin and Comparator Activity Against Select Enterobacteriaceae, Pseudomonas, and Enterococcus Urinary Tract InfectionDocument11 pagesFosfomycin and Comparator Activity Against Select Enterobacteriaceae, Pseudomonas, and Enterococcus Urinary Tract InfectionNawwal NaeemNo ratings yet
- Antimicrobial Resistances: The World's Next Pandemic On The WayDocument11 pagesAntimicrobial Resistances: The World's Next Pandemic On The WayIAEME PublicationNo ratings yet
- J. - Antimicrob. - Chemother. 2013 Vandecasteele 743 8Document6 pagesJ. - Antimicrob. - Chemother. 2013 Vandecasteele 743 8Natalia MoranNo ratings yet
- Vancomycin DemystifiedDocument4 pagesVancomycin DemystifiedTim KNo ratings yet
- The Use of Vancomycin With Its Therapeutic and Adverse Effects: A ReviewDocument11 pagesThe Use of Vancomycin With Its Therapeutic and Adverse Effects: A Reviewdea yuniarNo ratings yet
- Red Blood Cell Membrane-Camouflaged Vancomycin and ChlorogenicDocument16 pagesRed Blood Cell Membrane-Camouflaged Vancomycin and ChlorogenicMuhammad Shehr YarNo ratings yet
- Mechanisms of Antimicrobial Resistance in ESKAPE PathogensDocument8 pagesMechanisms of Antimicrobial Resistance in ESKAPE PathogensCinthia MadeiraNo ratings yet
- VREAbreifreviewDocument6 pagesVREAbreifreviewVini KastilaNo ratings yet
- Correspondence: Broad-And Narrow-Spectrum Antibiotics: A Different ApproachDocument2 pagesCorrespondence: Broad-And Narrow-Spectrum Antibiotics: A Different ApproachJulio Andro ArtamulandikaNo ratings yet
- s12879 019 4536 8Document11 pagess12879 019 4536 8mariamaNo ratings yet
- Ivermectin Potential Anticancer DrugDocument11 pagesIvermectin Potential Anticancer DrugJesus Bueno MorenoNo ratings yet
- Assessment of Susceptibility To Five Common Antibiotics and Their Resistance Pattern in Clinical Enterococcus IsolatesDocument10 pagesAssessment of Susceptibility To Five Common Antibiotics and Their Resistance Pattern in Clinical Enterococcus IsolatesIsidora NikolicNo ratings yet
- Journal of Advanced Research: Yanguang Cong, Sijin Yang, Xiancai RaoDocument8 pagesJournal of Advanced Research: Yanguang Cong, Sijin Yang, Xiancai RaoDebdeep MondalNo ratings yet
- br_16_1_1490_PDFDocument6 pagesbr_16_1_1490_PDFMarketable StudioNo ratings yet
- 2019 First-In-Human, Randomized, Double-Blind Clinical Trial of Differentially Adjuvanted PAMVAC, A Vaccine Candidate To Prevent Pregnancy-Associated MalariaDocument8 pages2019 First-In-Human, Randomized, Double-Blind Clinical Trial of Differentially Adjuvanted PAMVAC, A Vaccine Candidate To Prevent Pregnancy-Associated MalariaSethawud ChaikitgosiyakulNo ratings yet
- Daptomycin: Discovery, Development and Perspectives: Ipsita ChakravartyDocument9 pagesDaptomycin: Discovery, Development and Perspectives: Ipsita ChakravartyIpsita ChakravartyNo ratings yet
- dst15011 917 921Document5 pagesdst15011 917 921mariam farragNo ratings yet
- Hrabak 2014Document15 pagesHrabak 2014Ignacio Velasquez MiñoNo ratings yet
- Ofac 034Document6 pagesOfac 034Imene FlNo ratings yet
- Correlation Between A Baumanii Resistance and Hospital Use of Meropenem 2021Document12 pagesCorrelation Between A Baumanii Resistance and Hospital Use of Meropenem 2021czikkely23No ratings yet
- Kausar Et Al 2021 A Review Mechanism of Action of Antiviral DrugsDocument14 pagesKausar Et Al 2021 A Review Mechanism of Action of Antiviral Drugssimon pierre GAHIMBARENo ratings yet
- Safdar Et Al 2004 in Vivo Pharmacodynamic Activity of DaptomycinDocument6 pagesSafdar Et Al 2004 in Vivo Pharmacodynamic Activity of DaptomycinduongnguyenNo ratings yet
- Journal 8Document7 pagesJournal 8margareth silaenNo ratings yet
- Colson 2016Document4 pagesColson 2016rabbanibreNo ratings yet
- EditorialDocument4 pagesEditorialDiegoSilvaNo ratings yet
- Mechanisms of Linezolid Resistance in Staphylococci and Enterococci Isolated From Two Teaching Hospitals in Shanghai, ChinaDocument15 pagesMechanisms of Linezolid Resistance in Staphylococci and Enterococci Isolated From Two Teaching Hospitals in Shanghai, Chinanadila oktaviaNo ratings yet
- 4-Quinolone Hybrids and Their Antibacterial ActivitiesDocument11 pages4-Quinolone Hybrids and Their Antibacterial ActivitiesKevin ArechigaNo ratings yet
- Guide To Infection ControlDocument9 pagesGuide To Infection ControlMita RefanitaNo ratings yet
- Highly Effective Against EsblDocument9 pagesHighly Effective Against Esblgaurav rawalNo ratings yet
- 5 E Coli in UTIDocument7 pages5 E Coli in UTIGhadah AlyousifNo ratings yet
- TargetsDocument14 pagesTargetsLYDA ZAMIR LUENGASNo ratings yet
- Once-Daily Plazomicin For Complicated Urinary Tract InfectionsDocument12 pagesOnce-Daily Plazomicin For Complicated Urinary Tract InfectionsharvardboyNo ratings yet
- Antimalarial Drugs: Modes of Action and Mechanisms of Parasite ResistanceDocument18 pagesAntimalarial Drugs: Modes of Action and Mechanisms of Parasite ResistancePuspa Lestari RyathNo ratings yet
- Pharmaceutical Sciences: A Review On Aminoglycoside AntibioticsDocument9 pagesPharmaceutical Sciences: A Review On Aminoglycoside AntibioticsiajpsNo ratings yet
- TX Carbapenem Resistent MoDocument11 pagesTX Carbapenem Resistent MoInes MendonçaNo ratings yet
- Kovachev 2014Document7 pagesKovachev 2014habibNo ratings yet
- AFDDiStefano RifamycinSV AACDocument7 pagesAFDDiStefano RifamycinSV AACAndrea Di StefanoNo ratings yet
- Peter Hawkey: The Enemy Within - Hospital-Acquired, Antibiotic-Resistant BacteriaDocument3 pagesPeter Hawkey: The Enemy Within - Hospital-Acquired, Antibiotic-Resistant BacteriaCoffee CojjeeNo ratings yet
- Dalbavancin in The Treatment of Complicated Skin and Soft-Tissue Infections: A ReviewDocument10 pagesDalbavancin in The Treatment of Complicated Skin and Soft-Tissue Infections: A ReviewDiana Giorgiana DajbocNo ratings yet
- 1 Zuo2018Document7 pages1 Zuo2018Marco PoloNo ratings yet
- Novel Molecular Target in Malaria Parasite ChemotherapyDocument25 pagesNovel Molecular Target in Malaria Parasite ChemotherapyTimothyNo ratings yet
- s13613 017 0296 ZDocument7 pagess13613 017 0296 ZElsiana LaurenciaNo ratings yet
- Optimizing Vancomycin Dosing Through Pharmacodynamic Assessment Targeting Area Under The Concentration-Time Curve/Minimum Inhibitory ConcentrationDocument15 pagesOptimizing Vancomycin Dosing Through Pharmacodynamic Assessment Targeting Area Under The Concentration-Time Curve/Minimum Inhibitory ConcentrationMarcel O.No ratings yet
- Ajo Vre 052017Document3 pagesAjo Vre 052017api-383596435No ratings yet
- Detection of Methicillin Resistance in Staphylococcus Species From Clinical SamplesDocument6 pagesDetection of Methicillin Resistance in Staphylococcus Species From Clinical SamplesOpenaccess Research paperNo ratings yet
- alm-34-1Document7 pagesalm-34-1VidaNo ratings yet
- Antimicrobial Resistance in Methicillin-ResistantDocument14 pagesAntimicrobial Resistance in Methicillin-ResistantAlex HernandezNo ratings yet
- Medical Competence: The Interplay Between Individual Ability and The Health Care EnvironmentDocument7 pagesMedical Competence: The Interplay Between Individual Ability and The Health Care EnvironmentOscar F RojasNo ratings yet
- A Trial of A Shorter Regimen For Rifampin-Resistant TuberculosisDocument13 pagesA Trial of A Shorter Regimen For Rifampin-Resistant TuberculosisDiana SeptiyantiNo ratings yet
- Evolucion de PieDocument1 pageEvolucion de PieOscar F RojasNo ratings yet
- Ten Tips On FeverDocument4 pagesTen Tips On FeverOscar F RojasNo ratings yet
- Diabetic Foot Ulcers and Their RecurrenceDocument9 pagesDiabetic Foot Ulcers and Their RecurrenceRoberto López MataNo ratings yet
- Calcific Aortic Stenosis - Time To Look More Closely at The Valve PDFDocument4 pagesCalcific Aortic Stenosis - Time To Look More Closely at The Valve PDFOscar F RojasNo ratings yet
- Actividades Profesionales ConfiablesDocument6 pagesActividades Profesionales ConfiablesOscar F RojasNo ratings yet
- Chronic Venous Disease: Review ArticleDocument11 pagesChronic Venous Disease: Review ArticleOscar F RojasNo ratings yet
- EPAS Como Evaluacion de CompetenciasDocument6 pagesEPAS Como Evaluacion de CompetenciasEdwin MonarNo ratings yet
- Vancomicina ResistenciaDocument12 pagesVancomicina ResistenciaOscar F RojasNo ratings yet
- Calcific Aortic Stenosis - Time To Look More Closely at The Valve PDFDocument4 pagesCalcific Aortic Stenosis - Time To Look More Closely at The Valve PDFOscar F RojasNo ratings yet
- Calcific Aortic Stenosis - Time To Look More Closely at The Valve PDFDocument4 pagesCalcific Aortic Stenosis - Time To Look More Closely at The Valve PDFOscar F RojasNo ratings yet
- Aortic Stenosis: Changing Disease Concepts: ReviewDocument11 pagesAortic Stenosis: Changing Disease Concepts: ReviewdiogoNo ratings yet
- Angina Pectoris in Patients With Aortic Stenosis and Normal Coronary Arteries - CirculationDocument24 pagesAngina Pectoris in Patients With Aortic Stenosis and Normal Coronary Arteries - CirculationOscar F RojasNo ratings yet
- Oritavancin: Mechanism of Action: SupplementarticleDocument6 pagesOritavancin: Mechanism of Action: SupplementarticleOscar F RojasNo ratings yet
- Linezolid Update. Stable in Vitro Activity Following More Than A Decade PDFDocument12 pagesLinezolid Update. Stable in Vitro Activity Following More Than A Decade PDFsilvia mendez riveraNo ratings yet
- Benjamin P. Howden, John K. Davies, Paul D. R. Johnson, Timothy P. Stinear, and M. Lindsay GraysonDocument41 pagesBenjamin P. Howden, John K. Davies, Paul D. R. Johnson, Timothy P. Stinear, and M. Lindsay GraysonFernando Martínez HiguerasNo ratings yet
- Best Clinical Practice: Red Blood Cell Transfusion in The Emergency DepartmentDocument11 pagesBest Clinical Practice: Red Blood Cell Transfusion in The Emergency DepartmentOscar F RojasNo ratings yet
- Acute Colonic Diverticulitis: Annals of Internal MedicineDocument16 pagesAcute Colonic Diverticulitis: Annals of Internal MedicineOscar F RojasNo ratings yet
- Arritmias en Miocarditis ViralDocument13 pagesArritmias en Miocarditis ViralOscar F RojasNo ratings yet
- Hap Vap Guidelines 2016 PDFDocument51 pagesHap Vap Guidelines 2016 PDFJerryEddyaPutraBoerNo ratings yet
- Aortic Stenosis, A Left Ventricular Disease Insights From Advanced ImagingDocument9 pagesAortic Stenosis, A Left Ventricular Disease Insights From Advanced ImagingOscar F RojasNo ratings yet
- Clinical1bst PDFDocument182 pagesClinical1bst PDFHabibNo ratings yet
- Quantifying Bacterial Tolerance with a New Experimental FrameworkDocument8 pagesQuantifying Bacterial Tolerance with a New Experimental FrameworkOscar F RojasNo ratings yet
- Use of Blood Products in The Critically Ill - UpToDateDocument13 pagesUse of Blood Products in The Critically Ill - UpToDateOscar F RojasNo ratings yet
- AntibioticsDocument7 pagesAntibioticsOscar F RojasNo ratings yet
- English ID 3 TB 2013 193pDocument193 pagesEnglish ID 3 TB 2013 193pXimenita Bel80% (10)
- Acutw Gastrrointestrinal BleedingDocument16 pagesAcutw Gastrrointestrinal BleedingOscar F RojasNo ratings yet
- Cases in Primary CareDocument60 pagesCases in Primary CareOscar F Rojas100% (1)
- DR - PARTHA - 2018 Clinical Microbiology II PDFDocument88 pagesDR - PARTHA - 2018 Clinical Microbiology II PDFNuzki YofandaNo ratings yet
- DIT Pharmacology MnemonicsDocument5 pagesDIT Pharmacology MnemonicsqiqizNo ratings yet
- Dispensing AnsDocument197 pagesDispensing AnsPrincess Ronsable100% (1)
- 1 s2.0 S0169409X16300552 MainDocument19 pages1 s2.0 S0169409X16300552 MainpavaniNo ratings yet
- Vancomycin: A Powerful Antibiotic for Gram-Positive InfectionsDocument13 pagesVancomycin: A Powerful Antibiotic for Gram-Positive Infectionsفاطمة عدنان حسينNo ratings yet
- Delafloxacin 4Document23 pagesDelafloxacin 4Muhammad RamzanNo ratings yet
- Lecture 10 Antibiotics & Antiinfectives-1Document39 pagesLecture 10 Antibiotics & Antiinfectives-1Dr Haris AwanNo ratings yet
- Microbiology Lecture - 04 AntibioticsDocument72 pagesMicrobiology Lecture - 04 AntibioticsChris Queiklin100% (1)
- Review of Antibiotic Use in PregnancyDocument11 pagesReview of Antibiotic Use in Pregnancynoname19191No ratings yet
- Neutropenic FeverDocument15 pagesNeutropenic Feverapi-3712326No ratings yet
- Pharmacology: By: Nerissa Cabañero Laiza PinedaDocument121 pagesPharmacology: By: Nerissa Cabañero Laiza PinedaJacq CalaycayNo ratings yet
- SHC Vancomycin Dosing GuideDocument10 pagesSHC Vancomycin Dosing GuideAlan SmithNo ratings yet
- Endophthalmitis Harry Flynn MD OEDocument31 pagesEndophthalmitis Harry Flynn MD OEMsai AhyarNo ratings yet
- Diabetic Foot Infections: by Scott Bergman, Pharm.D., BCPS-AQ ID and Punit J. Shah, Pharm.D., BCPSDocument20 pagesDiabetic Foot Infections: by Scott Bergman, Pharm.D., BCPS-AQ ID and Punit J. Shah, Pharm.D., BCPSPrashant LokhandeNo ratings yet
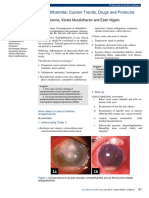
- Endophthalmitis: Current Trends, Drugs and Protocols: Aditya Verma, Vinata Muralidharan and Eesh NigamDocument10 pagesEndophthalmitis: Current Trends, Drugs and Protocols: Aditya Verma, Vinata Muralidharan and Eesh NigamHerman Kurt Ludvik100% (1)
- NICORANDIL PP Degradasi Nicotinamide Ada Rumus SIPROFLOKSASIN SENT1Document22 pagesNICORANDIL PP Degradasi Nicotinamide Ada Rumus SIPROFLOKSASIN SENT1Karillda Agnoin WandhaNo ratings yet
- Vancomycin Protocol: A Protocol To Guide The Dosing and Monitoring of Vancomycin in AdultsDocument5 pagesVancomycin Protocol: A Protocol To Guide The Dosing and Monitoring of Vancomycin in AdultsSaadia MahmoodNo ratings yet
- Share Pharmacology Book 1Document101 pagesShare Pharmacology Book 1Jmcle AhmedNo ratings yet
- Longitudinal Clinical Evaluation of Antibiotic Use Among Patients With InfectionDocument11 pagesLongitudinal Clinical Evaluation of Antibiotic Use Among Patients With InfectionMITA RESTINIA UINJKTNo ratings yet
- Fever in Children With ChemoteraphyDocument23 pagesFever in Children With ChemoteraphyImanuel Far-FarNo ratings yet
- دوائيات ن- lec22-ReducedDocument24 pagesدوائيات ن- lec22-Reducedma22mdmmNo ratings yet
- ? PHARMA 3. Anti InfectivesDocument130 pages? PHARMA 3. Anti InfectivesMiguel Luis NavarreteNo ratings yet
- PANCE Antimicrobial Review BlueprintDocument40 pagesPANCE Antimicrobial Review BlueprintFlora LawrenceNo ratings yet
- CephalosporinsDocument19 pagesCephalosporinsAliImadAlKhasakiNo ratings yet
- Drug ChartDocument8 pagesDrug Chartstudentalwaysstudy100% (1)
- VancomycinDocument2 pagesVancomycinRye AnchNo ratings yet
- VancomycinDocument1 pageVancomycinE100% (2)
- Daptomycin: Discovery, Development and Perspectives: Ipsita ChakravartyDocument9 pagesDaptomycin: Discovery, Development and Perspectives: Ipsita ChakravartyIpsita ChakravartyNo ratings yet
- PDF PedirefcardDocument4 pagesPDF PedirefcardRaditya Indah TofaniNo ratings yet
- Antibiotics Use in Orthopaedic Surgery An OverviewDocument7 pagesAntibiotics Use in Orthopaedic Surgery An OverviewnjmdrNo ratings yet