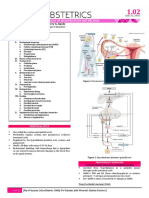
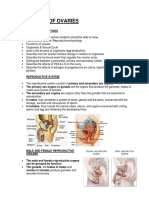
You might also like
- Vulvovaginal InfectionDocument37 pagesVulvovaginal Infectionmadmax500No ratings yet
- Central Limit Theorem WorksheetDocument2 pagesCentral Limit Theorem WorksheetChris TianNo ratings yet
- Physiology of OvulationDocument7 pagesPhysiology of OvulationAza Patullah ZaiNo ratings yet
- Hormone Action in The Mammary Gland - C Brisken and B O Malley 2010Document15 pagesHormone Action in The Mammary Gland - C Brisken and B O Malley 2010tostonasNo ratings yet
- The Female Reproductive System: Paul F. Terranova, PH.DDocument17 pagesThe Female Reproductive System: Paul F. Terranova, PH.DMekuriya BeregaNo ratings yet
- JurnalDocument4 pagesJurnalS1kebidanan Semester 3No ratings yet
- Tog 12665Document13 pagesTog 12665saeed hasan saeed100% (1)
- Stages of Puberty - Sundhed - DKDocument6 pagesStages of Puberty - Sundhed - DKPavel BerlinschiNo ratings yet
- Endocrinology of The MenopauseDocument12 pagesEndocrinology of The MenopauseGina Morales FerrerNo ratings yet
- Fisiologi HaidDocument66 pagesFisiologi HaidRizna AriyaniNo ratings yet
- Ciclo Estral de La CerdaDocument8 pagesCiclo Estral de La CerdaAndrea GranadosNo ratings yet
- Female Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeDocument56 pagesFemale Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeVFJulianNo ratings yet
- 10-2 Ambeguia, King Rainier C. (LAS1 W 1 3rd Grading)Document4 pages10-2 Ambeguia, King Rainier C. (LAS1 W 1 3rd Grading)Elijah Ronnie C. AmbeguiaNo ratings yet
- Normal Menstrual Cycle The ProcessDocument6 pagesNormal Menstrual Cycle The ProcessBrett StevensonNo ratings yet
- It Takes Two To TangoDocument3 pagesIt Takes Two To Tangoaftab khanNo ratings yet
- Periconceptional Period PDFDocument57 pagesPericonceptional Period PDFJamesNo ratings yet
- Sessions 1 and 2 - 2023 FinalDocument34 pagesSessions 1 and 2 - 2023 FinalChen ZhonghaoNo ratings yet
- MODULE 1-Lesson 2-3Document11 pagesMODULE 1-Lesson 2-3JOHN BRAINARD PEJONo ratings yet
- Cold Spring Harb Perspect Biol-2010-Brisken-a003178Document17 pagesCold Spring Harb Perspect Biol-2010-Brisken-a003178wi bowoNo ratings yet
- Female HormoneDocument43 pagesFemale Hormoneامجد حسين جواد كاظمNo ratings yet
- Female Physiology Before Pregnancy and The Female HormonesDocument3 pagesFemale Physiology Before Pregnancy and The Female HormonesspringdingNo ratings yet
- Disorders of The Female Reproductive System PDFDocument4 pagesDisorders of The Female Reproductive System PDFSeff CausapinNo ratings yet
- Handout On ReproDocument4 pagesHandout On ReproJoe JosephNo ratings yet
- Understanding Human Sexuality 13th Edition Hyde Solutions Manual 1Document21 pagesUnderstanding Human Sexuality 13th Edition Hyde Solutions Manual 1Larry Kershaw100% (41)
- Understanding Human Sexuality 13Th Edition Hyde Solutions Manual Full Chapter PDFDocument36 pagesUnderstanding Human Sexuality 13Th Edition Hyde Solutions Manual Full Chapter PDFcharlotte.gammon593100% (11)
- Follicular PhaseDocument9 pagesFollicular PhaseDista ArifanyNo ratings yet
- Female PhysiologyDocument10 pagesFemale PhysiologyMZ LimNo ratings yet
- UntitledDocument8 pagesUntitledPJ Manuel SuerteNo ratings yet
- Oxytocin, ADH Lecture For 2nd Year MBBS by DR Waeem KausarDocument28 pagesOxytocin, ADH Lecture For 2nd Year MBBS by DR Waeem KausarIMDCBiochemNo ratings yet
- Hormones and the Fetus: Volume 1: Production, Concentration and Metabolism During PregnancyFrom EverandHormones and the Fetus: Volume 1: Production, Concentration and Metabolism During PregnancyNo ratings yet
- The Menstrual Cycle Ob - Gyn SecretsDocument4 pagesThe Menstrual Cycle Ob - Gyn Secretsleonardotorres1No ratings yet
- Neuronendocrine Control of The Onset of Puberty 2015 PlantDocument16 pagesNeuronendocrine Control of The Onset of Puberty 2015 PlantCarlos MartínezNo ratings yet
- OB 1 - 1.02 Physiology of Menstruation and Decidua PDFDocument11 pagesOB 1 - 1.02 Physiology of Menstruation and Decidua PDFMDreamerNo ratings yet
- Female Physiology 1Document2 pagesFemale Physiology 1JayricDepalobosNo ratings yet
- Reproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, OrlandoDocument10 pagesReproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, Orlandokuku411No ratings yet
- Mammalian Reproductive SystemDocument50 pagesMammalian Reproductive SystemLeena MuniandyNo ratings yet
- Hyperpituitarism Group e Bsn3Document17 pagesHyperpituitarism Group e Bsn3Mari IllustriousNo ratings yet
- The Normal Menstrual CycleDocument12 pagesThe Normal Menstrual CycleDheyaa A. SabahNo ratings yet
- Functions of Ovaries: Learning ObjectivesDocument9 pagesFunctions of Ovaries: Learning ObjectivesUloko ChristopherNo ratings yet
- Fertilization Reflection PaperDocument2 pagesFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- Basic Concepts of SexDocument2 pagesBasic Concepts of SexEricka Mae FedereNo ratings yet
- Reproductive SystemDocument30 pagesReproductive SystemmfaizchejamriNo ratings yet
- Hormone-A Chemical Substance Produced in The Body That Controls and RegulatesDocument2 pagesHormone-A Chemical Substance Produced in The Body That Controls and RegulatesJohn Cyrus DizonNo ratings yet
- EndocrinologyDocument7 pagesEndocrinologyVijith.V.kumarNo ratings yet
- Nzary El 3maliDocument13 pagesNzary El 3maliGana KhaledNo ratings yet
- Fisiologi Siklus HaidDocument70 pagesFisiologi Siklus HaidsilviNo ratings yet
- Menstrual Physiology, Amenorrhea: Page 1 of 5Document5 pagesMenstrual Physiology, Amenorrhea: Page 1 of 5seth10No ratings yet
- Puberty, Menstrual CycleDocument9 pagesPuberty, Menstrual CycleHalo Ini JeskryNo ratings yet
- Problem No.1Document34 pagesProblem No.1ayesha bgNo ratings yet
- The Female Reproductive System - 1Document9 pagesThe Female Reproductive System - 1Sherida GibbsNo ratings yet
- Javern M. Supnet RNDocument16 pagesJavern M. Supnet RNGenghis SupnetNo ratings yet
- Definition of EmbryologyDocument12 pagesDefinition of EmbryologyOmar BasimNo ratings yet
- Fact Sheet 1 (Menstrual Cycle)Document2 pagesFact Sheet 1 (Menstrual Cycle)BeaNo ratings yet
- Bio12 c10 10 7Document8 pagesBio12 c10 10 7Eric McMullenNo ratings yet
- Cow EstrusDocument4 pagesCow EstrusMageja TatendaNo ratings yet
- Female Reproductive SystemDocument3 pagesFemale Reproductive SystemJayricDepalobosNo ratings yet
- NIH Public Access: Hormonal Regulation of Female ReproductionDocument9 pagesNIH Public Access: Hormonal Regulation of Female ReproductionReski Rahmadani BakriNo ratings yet
- Menstruation & OvulationDocument42 pagesMenstruation & Ovulationneilesh300No ratings yet
- RS-1 K5-K6 Hormon Repro Pria Dan Wanita, EditDocument57 pagesRS-1 K5-K6 Hormon Repro Pria Dan Wanita, EditfelixNo ratings yet
- Physiology of The Menstrual Cycle 2Document22 pagesPhysiology of The Menstrual Cycle 2anoy judeNo ratings yet
- Frontiers in Neuroendocrinology: Tony M. PlantDocument16 pagesFrontiers in Neuroendocrinology: Tony M. PlantRUBY JOJOANo ratings yet
- Billings Method in The Service of Knowledge, Love and Science Aurora SaporosiDocument18 pagesBillings Method in The Service of Knowledge, Love and Science Aurora SaporosiCaro Lascano CorreaNo ratings yet
- Regulating Glucagon Secretion: Somatostatin in The SpotlightDocument3 pagesRegulating Glucagon Secretion: Somatostatin in The SpotlightDhikaNo ratings yet
- Kasus2Blefarokonjungtivitis BlokSSS Tingkat1Document146 pagesKasus2Blefarokonjungtivitis BlokSSS Tingkat1DhikaNo ratings yet
- Kasus2Blefarokonjungtivitis BlokSSS Tingkat1Document146 pagesKasus2Blefarokonjungtivitis BlokSSS Tingkat1DhikaNo ratings yet
- The Trusted Source For Healthcare Information And: Continuing EducationDocument19 pagesThe Trusted Source For Healthcare Information And: Continuing EducationDhikaNo ratings yet
- Glucagon Receptor Signaling and Lipid MetabolismDocument11 pagesGlucagon Receptor Signaling and Lipid MetabolismDhikaNo ratings yet
- Hormonal Characteristics of The Throughout Reproductive LifeDocument8 pagesHormonal Characteristics of The Throughout Reproductive LifeDhikaNo ratings yet
- Pelvis and Perineum - Anatomy, Vessels, Nerves - KenhubDocument11 pagesPelvis and Perineum - Anatomy, Vessels, Nerves - KenhubDhika100% (1)
- Teaching Sexuality To ChildrenDocument6 pagesTeaching Sexuality To ChildrenNguyễn Ngọc Quỳnh NhưNo ratings yet
- Health and Population IndicatorsDocument68 pagesHealth and Population IndicatorsMarites MaylanonNo ratings yet
- NCM 107 MidtermDocument34 pagesNCM 107 MidtermChiaraNo ratings yet
- Part 4 MAPEH PDFDocument5 pagesPart 4 MAPEH PDFGemma AmazonaNo ratings yet
- Ectopic Pregnancy For Med Students ExplainedDocument2 pagesEctopic Pregnancy For Med Students Explainedmartin magdenoskiNo ratings yet
- Pelvic Pain, Food Allergies, Thyroid Disease, Mpox, Chronic Fatigue Syndrome, Alloimmunization (AAFP 2023)Document1 pagePelvic Pain, Food Allergies, Thyroid Disease, Mpox, Chronic Fatigue Syndrome, Alloimmunization (AAFP 2023)Breno Resende Rodrigues CunhaNo ratings yet
- Endocrine Gland - Wikipedia, The Free EncyclopediaDocument5 pagesEndocrine Gland - Wikipedia, The Free EncyclopediakbhattacNo ratings yet
- June 2012 (v3) QP - Paper 3 CIE Biology IGCSE PDFDocument24 pagesJune 2012 (v3) QP - Paper 3 CIE Biology IGCSE PDFAli AshrafNo ratings yet
- NDTDocument1 pageNDTAngela CancinoNo ratings yet
- Handout - Coercion and EugenicsDocument6 pagesHandout - Coercion and EugenicsAyah AbusafiaNo ratings yet
- The Socio Cultural Factors Affecting The Use of Family Planning Among Rural Women in Gwagwalada Area Council, Fct.Document5 pagesThe Socio Cultural Factors Affecting The Use of Family Planning Among Rural Women in Gwagwalada Area Council, Fct.David SamNo ratings yet
- MCQ Physiology Mid - Sarah Alsalman PDFDocument3 pagesMCQ Physiology Mid - Sarah Alsalman PDFNay Chi Thant Zin100% (1)
- PCPNDT ActDocument25 pagesPCPNDT ActHema Anand100% (1)
- AP Maternal Health - District Review - 17!03!2021Document58 pagesAP Maternal Health - District Review - 17!03!2021primary health centre dowleswaramNo ratings yet
- Water BirthDocument9 pagesWater Birthannu panchalNo ratings yet
- Artikel Hubungan Usia Kehamilan Dengan Kejadian Stretch Mark Pada Ibu HamilDocument9 pagesArtikel Hubungan Usia Kehamilan Dengan Kejadian Stretch Mark Pada Ibu Hamiljunaidi alimunarNo ratings yet
- Stability Continence and BreathingDocument16 pagesStability Continence and Breathingmemoona kaleemNo ratings yet
- Nursing Diagnosis Nursing Goals Nursing Outcome: (Goal Attainable Within The Shift) (With Rationale & Source)Document2 pagesNursing Diagnosis Nursing Goals Nursing Outcome: (Goal Attainable Within The Shift) (With Rationale & Source)merryNo ratings yet
- Vacuum DeliveryDocument31 pagesVacuum DeliveryAparna LaxmanNo ratings yet
- Life Sciences Revision 2022 DocumentDocument89 pagesLife Sciences Revision 2022 Documentabenathichagi2010No ratings yet
- 50 Human Studies, in Utero, Con - Jim WestDocument19 pages50 Human Studies, in Utero, Con - Jim WestAdrian BoboceaNo ratings yet
- Chapter 2 Review of Related Literature ADocument14 pagesChapter 2 Review of Related Literature ANicole CariasNo ratings yet
- Safety and Health at Work: Kyung-Taek RimDocument8 pagesSafety and Health at Work: Kyung-Taek RimAkbarNo ratings yet
- Experience of Mental Health in Women With Polycystic Ovary Syndrome A Descriptive Phenomenological StudyDocument11 pagesExperience of Mental Health in Women With Polycystic Ovary Syndrome A Descriptive Phenomenological StudyMelany AntounNo ratings yet
- Bibliography of AbortionDocument10 pagesBibliography of AbortioncherubrockNo ratings yet
- Learning Activity Sheet (Las) : Department of EducationDocument15 pagesLearning Activity Sheet (Las) : Department of EducationJoeffrie G. Costales100% (1)
- Adolescence and Puberty PDFDocument318 pagesAdolescence and Puberty PDFjorge torresNo ratings yet
- Tami Taylor, Mother To All: Abortion On Friday Night LightsDocument23 pagesTami Taylor, Mother To All: Abortion On Friday Night LightsRenee PowersNo ratings yet