You might also like
- Pulmonary Functional Imaging: Basics and Clinical ApplicationsFrom EverandPulmonary Functional Imaging: Basics and Clinical ApplicationsYoshiharu OhnoNo ratings yet
- Editorial The Future of Sensors and Instrumentation For Human Breath AnalysisDocument4 pagesEditorial The Future of Sensors and Instrumentation For Human Breath AnalysisLukman MahdiNo ratings yet
- Biomarkers in COPD ExacerbationsDocument12 pagesBiomarkers in COPD ExacerbationsLinto JohnNo ratings yet
- Jurnal BiomedikDocument10 pagesJurnal BiomedikneaNo ratings yet
- Applsci 09 02842 PDFDocument13 pagesApplsci 09 02842 PDFFelicia SutarliNo ratings yet
- 2004 Clinica Chimica ActaDocument15 pages2004 Clinica Chimica ActaDina MansourNo ratings yet
- Human Exhaled Breath Analysis: CME ReviewDocument6 pagesHuman Exhaled Breath Analysis: CME ReviewJulieta SolariNo ratings yet
- Exhaled Breath Analysis in The Differentiation of Pneumonia From Acute Pulmonary OedemaDocument7 pagesExhaled Breath Analysis in The Differentiation of Pneumonia From Acute Pulmonary OedemaAkka RakaNo ratings yet
- Breath Analysis: Trends in Techniques and Clinical ApplicationsDocument6 pagesBreath Analysis: Trends in Techniques and Clinical ApplicationsDina MansourNo ratings yet
- Exhaled Breath Analysis For Lung CancerDocument12 pagesExhaled Breath Analysis For Lung CancerIkaPKNo ratings yet
- Unit 7 Blood PressureDocument11 pagesUnit 7 Blood PressureQ O R Y A N INo ratings yet
- Analysis of Volatile Organic Compounds From Deep Airway in The Lung Through Intubation SamplingDocument12 pagesAnalysis of Volatile Organic Compounds From Deep Airway in The Lung Through Intubation SamplingmoubarginNo ratings yet
- 2005 Determination of Acetone in Breath Original ResearchDocument11 pages2005 Determination of Acetone in Breath Original ResearchDina MansourNo ratings yet
- Brock 2017Document10 pagesBrock 2017Carlos Daniel AliagaNo ratings yet
- Exhaled Breath PaperDocument16 pagesExhaled Breath PaperCristhian Manuel Durán AcevedoNo ratings yet
- Optoelectronic PlethysmographyDocument15 pagesOptoelectronic PlethysmographygothicmetalspiritNo ratings yet
- PATH Recommendations - EditedDocument2 pagesPATH Recommendations - EditedJohnNo ratings yet
- Capnography in DogsDocument14 pagesCapnography in DogswalczakcNo ratings yet
- 2006 129 1344-1348 Christopher E. Brightling: Clinical Applications of Induced SputumDocument7 pages2006 129 1344-1348 Christopher E. Brightling: Clinical Applications of Induced Sputumjoal510No ratings yet
- Editorials: Assessing The Dose of Supplemental Oxygen: Let Us Compare MethodologiesDocument2 pagesEditorials: Assessing The Dose of Supplemental Oxygen: Let Us Compare MethodologiesJafar JilaniNo ratings yet
- Lung Function and Peak Oxygen Uptake in Chronic Obstructive Pulmonary Disease Phenotypes With and Without EmphysemaDocument12 pagesLung Function and Peak Oxygen Uptake in Chronic Obstructive Pulmonary Disease Phenotypes With and Without EmphysemaAndreea MoalesNo ratings yet
- Comparison of The Efectiveness of High-Fow Nasal Oxygen vs. Standard Facemask Oxygenation For Pre - and Apneic Oxygenation During Anesthesia InductionDocument1 pageComparison of The Efectiveness of High-Fow Nasal Oxygen vs. Standard Facemask Oxygenation For Pre - and Apneic Oxygenation During Anesthesia InductionJodhy MaulanaNo ratings yet
- Exhaled 8-Isoprostane As An Biomarker of Lung Oxidative Stress in Patients With COPD and Healthy SmokersDocument3 pagesExhaled 8-Isoprostane As An Biomarker of Lung Oxidative Stress in Patients With COPD and Healthy SmokersAnthonyJohanNo ratings yet
- An Adverse Outcome Pathway For Lung Surfactant Functio - 2021 - Current ResearchDocument12 pagesAn Adverse Outcome Pathway For Lung Surfactant Functio - 2021 - Current ResearchAadityarajsinh GohilNo ratings yet
- Jurnal TuberkulosisDocument12 pagesJurnal TuberkulosisviviavisaNo ratings yet
- Bmri2016 3068467Document5 pagesBmri2016 3068467atika sgrtNo ratings yet
- Ijms 13 13894Document17 pagesIjms 13 13894osama mosaNo ratings yet
- Biomarker Copd PDFDocument35 pagesBiomarker Copd PDFManoj KumarNo ratings yet
- Anestesiologi EmergencyDocument12 pagesAnestesiologi EmergencyJodhy MaulanaNo ratings yet
- Mulyadi Jurnal5Document5 pagesMulyadi Jurnal5amelia tantriNo ratings yet
- Pan 14208Document23 pagesPan 14208rizki romadaniNo ratings yet
- EmfizemDocument4 pagesEmfizemJelenaJankovNo ratings yet
- Pulmonary Function TestDocument3 pagesPulmonary Function Testhm3398No ratings yet
- Journal of Theoretical Biology: Angela Reynolds, G. Bard Ermentrout, Gilles ClermontDocument13 pagesJournal of Theoretical Biology: Angela Reynolds, G. Bard Ermentrout, Gilles ClermontstiggofthedumpNo ratings yet
- To Study The Effect of Buteyko Breathing Technique in Patients With Obstructive Airway DiseaseDocument15 pagesTo Study The Effect of Buteyko Breathing Technique in Patients With Obstructive Airway DiseaseGerry KrikhoffNo ratings yet
- Pulse Oximetry as an Instrumental Tool for Dysphagia EvaluationDocument8 pagesPulse Oximetry as an Instrumental Tool for Dysphagia EvaluationGabriel MaldonadoNo ratings yet
- Respiration - Electronic NoseDocument13 pagesRespiration - Electronic NoseCristhian Manuel Durán AcevedoNo ratings yet
- Sauca (M) I6157820 BSC Thesis 2020Document33 pagesSauca (M) I6157820 BSC Thesis 2020marius saucaNo ratings yet
- Peel (VOCs Asthma SR) 2020Document18 pagesPeel (VOCs Asthma SR) 2020Luis Taborda BarataNo ratings yet
- Measuring Lung Capacity in Men and WomenDocument11 pagesMeasuring Lung Capacity in Men and WomenFatmawatiNo ratings yet
- Asthma - ArticleDocument10 pagesAsthma - ArticleMichael CaldeiraNo ratings yet
- 2015 - Gaspard - Aerosol Sampling Using An Electrostatic Precipitator Integrated With A Microfluidic Interface - PaperDocument9 pages2015 - Gaspard - Aerosol Sampling Using An Electrostatic Precipitator Integrated With A Microfluidic Interface - PaperNgô Đình NhânNo ratings yet
- 744 FullDocument5 pages744 FullBogdan NechitaNo ratings yet
- Nitrous Oxide Diffusion and The Second Gas Effect.26Document7 pagesNitrous Oxide Diffusion and The Second Gas Effect.26Syukron AmrullahNo ratings yet
- CHEST Nov 2005Document183 pagesCHEST Nov 2005rizjkuramaNo ratings yet
- Exhaled Single-Breath Nitric Oxide Measurements Are Reproducible, Repeatable and Reflect Levels of Nitric Oxide Found in The Lower AirwaysDocument6 pagesExhaled Single-Breath Nitric Oxide Measurements Are Reproducible, Repeatable and Reflect Levels of Nitric Oxide Found in The Lower AirwaysDennis CobbNo ratings yet
- Medi 99 E23646Document8 pagesMedi 99 E23646panumatkhamphiranonNo ratings yet
- Pulmonary Pharmacology & TherapeuticsDocument10 pagesPulmonary Pharmacology & TherapeuticsJoko Arif KurniawanNo ratings yet
- Ambulatory Pulse Oximetry Monitoring in Japanese COPD Outpatients Not Receiving Oxygen TherapyDocument8 pagesAmbulatory Pulse Oximetry Monitoring in Japanese COPD Outpatients Not Receiving Oxygen Therapynurharyanti darmaningtyasNo ratings yet
- RPT 14 03Document126 pagesRPT 14 03CHRISTOS KALOUDISNo ratings yet
- Unraveling The Pathophysiology of The Asthma-COPD Overlap SyndromeDocument8 pagesUnraveling The Pathophysiology of The Asthma-COPD Overlap SyndromeRadhianie DjanNo ratings yet
- Analysis of Sputum Markers in The Evaluation of Lung Inflammation and Functional Impairment in Symptomatic Smokers and COPD PatientsDocument10 pagesAnalysis of Sputum Markers in The Evaluation of Lung Inflammation and Functional Impairment in Symptomatic Smokers and COPD PatientsAndreea MoalesNo ratings yet
- 2018 Article 654Document6 pages2018 Article 654fiqhiNo ratings yet
- Proteome Profiling of Lung Tissues in ChronicDocument8 pagesProteome Profiling of Lung Tissues in ChronicAndreea MoalesNo ratings yet
- Adenosine 5 Monophosphate Increases Levels of Leukotriene - 2004 - RespiratoryDocument5 pagesAdenosine 5 Monophosphate Increases Levels of Leukotriene - 2004 - RespiratoryIon CorbuNo ratings yet
- Ozone As A Modulator of The Immune SystemDocument10 pagesOzone As A Modulator of The Immune Systemweb3351No ratings yet
- DITIADocument5 pagesDITIADian PertiwiNo ratings yet
- Markers of Pulmonary Oxygen Toxicity in Hyperbaric Oxygen Therapy Using Exhaled Breath AnalysisDocument9 pagesMarkers of Pulmonary Oxygen Toxicity in Hyperbaric Oxygen Therapy Using Exhaled Breath AnalysisThijs WingelaarNo ratings yet
- Noninvasive Methods for Assessing Airway Inflammation and Structural Changes in Asthma and COPDDocument11 pagesNoninvasive Methods for Assessing Airway Inflammation and Structural Changes in Asthma and COPDAndreea MoalesNo ratings yet
- Altered Lung Biology of Healthy NeverDocument10 pagesAltered Lung Biology of Healthy NeverRobby RanidoNo ratings yet
- Eeg Ahora SiDocument20 pagesEeg Ahora SiLOS ANDES VIDRERIANo ratings yet
- Abg Quiz in Class With Answers 1Document3 pagesAbg Quiz in Class With Answers 1Janae TaylorNo ratings yet
- Acid Base Tutorial - University of ConnecticutDocument47 pagesAcid Base Tutorial - University of ConnecticutClaudioGutierrezFrausto80% (5)
- CHF: Exercise Therapy for Heart Failure ManagementDocument20 pagesCHF: Exercise Therapy for Heart Failure ManagementJoeNo ratings yet
- First Nursing DiagnosisDocument3 pagesFirst Nursing DiagnosisDarlene Joy FullaNo ratings yet
- Risk For AspirationDocument2 pagesRisk For AspirationErica FabrigasNo ratings yet
- Pediatric Sepsis Case Study Manuscript 1Document37 pagesPediatric Sepsis Case Study Manuscript 1Denise Gabrielle GutierrezNo ratings yet
- Tortora 13e Glossary 2012Document30 pagesTortora 13e Glossary 2012Shweta KhannaNo ratings yet
- Endocrine System: 3.6 Functions, Definitions & VocabularyDocument20 pagesEndocrine System: 3.6 Functions, Definitions & VocabularyD AngelaNo ratings yet
- Photosynthesis: Rafia Urooj Saman 2020-Ag-937Document18 pagesPhotosynthesis: Rafia Urooj Saman 2020-Ag-937Rafya ShabbirNo ratings yet
- The Rose That Grew From Concrete by Tupac ShakurDocument5 pagesThe Rose That Grew From Concrete by Tupac ShakurTiffany HorsfordNo ratings yet
- Article in Press: SarcopeniaDocument6 pagesArticle in Press: SarcopeniaLucas CardosoNo ratings yet
- NCA - CVA InfarctDocument126 pagesNCA - CVA InfarctRosaree Mae PantojaNo ratings yet
- BIOLOGY- HOMEOSTASIS REGULATIONDocument16 pagesBIOLOGY- HOMEOSTASIS REGULATIONJuliana NorNo ratings yet
- The Phoenix Protocol Dry Fasting For Rapid Healing and Radical Life Extension (August Dunning) (Z-Library)Document124 pagesThe Phoenix Protocol Dry Fasting For Rapid Healing and Radical Life Extension (August Dunning) (Z-Library)amitlkoyogaNo ratings yet
- Emergency Cardiovascular Life SupportDocument88 pagesEmergency Cardiovascular Life SupportMARK JOSHUA CRUZNo ratings yet
- Handbook of Pediatric Neurological Emergency Book NEWDocument174 pagesHandbook of Pediatric Neurological Emergency Book NEWAyidhNo ratings yet
- ELEC4810: Introduction To Biosensors and Bioinstrumentation: Lecture Notes - Set #9Document58 pagesELEC4810: Introduction To Biosensors and Bioinstrumentation: Lecture Notes - Set #9Kwan ChanNo ratings yet
- Exercise 15 Cardiovascular SystDocument6 pagesExercise 15 Cardiovascular Systholie madroneroNo ratings yet
- Nervous System - All NotesDocument6 pagesNervous System - All NotesJNo ratings yet
- Case Study 3 (Botulism)Document1 pageCase Study 3 (Botulism)Fariz LukmanNo ratings yet
- Sleep Health Reciprocal Regulation of Sleep and Innate ImmunityDocument27 pagesSleep Health Reciprocal Regulation of Sleep and Innate ImmunityPolly MollyNo ratings yet
- Initial Assessment and Management of TraumaDocument40 pagesInitial Assessment and Management of Traumayunita murfhiNo ratings yet
- Ventilation and PerfusionDocument2 pagesVentilation and PerfusionJayricDepalobosNo ratings yet
- Living with Rare IST Heart ConditionDocument4 pagesLiving with Rare IST Heart ConditionMartin huntNo ratings yet
- JawapanKeseluruhan 2-FlattenedDocument70 pagesJawapanKeseluruhan 2-FlattenedFikri RaisNo ratings yet
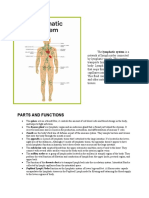
- Parts and FunctionsDocument4 pagesParts and FunctionsGerwin SilvestreNo ratings yet
- AR06894-Vigileo UserManual 1LRDocument31 pagesAR06894-Vigileo UserManual 1LRsec.ivbNo ratings yet
- Presentation on Guillain-Barre SyndromeDocument12 pagesPresentation on Guillain-Barre Syndromepandem soniyaNo ratings yet
- Biology Worksheet No. 2 9bDocument11 pagesBiology Worksheet No. 2 9bAli AshrafNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (16)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- CBT Strategies: CBT Strategies for Overcoming Panic, Fear, Depression, Anxiety, Worry, and AngerFrom EverandCBT Strategies: CBT Strategies for Overcoming Panic, Fear, Depression, Anxiety, Worry, and AngerNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- How To Understand Your Anger For Better Mental Health: How To Control Emotions And Eliminate Stress In Your LifeFrom EverandHow To Understand Your Anger For Better Mental Health: How To Control Emotions And Eliminate Stress In Your LifeNo ratings yet
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Inmersion Into The Shadown Effective Method Of Dark Psychology: How To Use The Best Persuasion Techniques To Achieve Your Best Goals And How To Protect Yourself From Being ManipulatedFrom EverandInmersion Into The Shadown Effective Method Of Dark Psychology: How To Use The Best Persuasion Techniques To Achieve Your Best Goals And How To Protect Yourself From Being ManipulatedRating: 4 out of 5 stars4/5 (1)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisFrom EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisRating: 5 out of 5 stars5/5 (8)