You might also like
- 000504242-Microwave Ephedrine From LpacDocument4 pages000504242-Microwave Ephedrine From LpacPaul BrownNo ratings yet
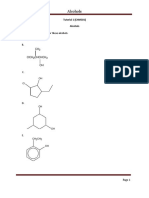
- Tutorial 1 - Alcohol PDFDocument5 pagesTutorial 1 - Alcohol PDFNurul Athirah JainiNo ratings yet
- Science: Quarter 4 - Module 6: Cycling of Materials in The EcosystemDocument20 pagesScience: Quarter 4 - Module 6: Cycling of Materials in The EcosystemCamille Sabal100% (1)
- Biology For Engineers Question Bank Module 1-3Document22 pagesBiology For Engineers Question Bank Module 1-3Edu techNo ratings yet
- Nakanishi 2014Document7 pagesNakanishi 2014Abhishek PareekNo ratings yet
- Organic Chemistry: Volume 23, Number 6 Ju N e 20, 1958Document160 pagesOrganic Chemistry: Volume 23, Number 6 Ju N e 20, 1958Alexander MeloNo ratings yet
- Example Report PDFDocument5 pagesExample Report PDFVn Dang100% (1)
- 3,3' Positions Was Achieved By: Dinitro-2,2'-BithiophenesDocument8 pages3,3' Positions Was Achieved By: Dinitro-2,2'-Bithiophenesshrujan9No ratings yet
- Indian BromacionDocument2 pagesIndian BromacionJosé Guadalupe García EstradaNo ratings yet
- Ibrahim Saad Abdel Ha Z, Elham Sayed Darwish and Fivian Farouk MahmoudDocument2 pagesIbrahim Saad Abdel Ha Z, Elham Sayed Darwish and Fivian Farouk MahmoudWalid EbaiedNo ratings yet
- 2-Amidinylindole-3-Carbaldehydes: Versatile Synthons For The Preparation of A-Carboline DerivativesDocument7 pages2-Amidinylindole-3-Carbaldehydes: Versatile Synthons For The Preparation of A-Carboline DerivativesWalid EbaiedNo ratings yet
- Temel2010 PDFDocument7 pagesTemel2010 PDFrommy agurto palaciosNo ratings yet
- Jo 000853Document8 pagesJo 000853natali.pfernandezNo ratings yet
- Nudelman 2004Document4 pagesNudelman 2004kongaradamuNo ratings yet
- Solubility - Mygdalin - Ethanol + WaterDocument17 pagesSolubility - Mygdalin - Ethanol + WaterDanielRicardoDelgadoNo ratings yet
- A Practical Synthesis of d-α-Terpineol via Markovnikov Addition of d-Limonene Using Trifluoroacetic AcidDocument2 pagesA Practical Synthesis of d-α-Terpineol via Markovnikov Addition of d-Limonene Using Trifluoroacetic AcidGreg HillNo ratings yet
- College of Arts and Sciences, Department of Chemistry, Drake UniversityDocument1 pageCollege of Arts and Sciences, Department of Chemistry, Drake UniversityBaloch SamNo ratings yet
- Iron-Catalyzed Cross-Coupling of Primary and Secondary Alkyl Halides With Aryl Grignard ReagentsDocument2 pagesIron-Catalyzed Cross-Coupling of Primary and Secondary Alkyl Halides With Aryl Grignard ReagentssamzamanNo ratings yet
- Amino Derivatives of Nitrochalcones. I. Synthesis, Structure Studies and Absorption SpectraDocument5 pagesAmino Derivatives of Nitrochalcones. I. Synthesis, Structure Studies and Absorption SpectraDiana ArizaNo ratings yet
- 10 11364@networkpolymer1996 27 218Document3 pages10 11364@networkpolymer1996 27 218Robert RicardoNo ratings yet
- Fibrino - Peptide: Fibrinogen-ThrombinDocument4 pagesFibrino - Peptide: Fibrinogen-ThrombinAziziNo ratings yet
- Eddy 2015Document3 pagesEddy 2015Gabriel FenteanyNo ratings yet
- Pyrolysis Polytetrafluoroethylene: A SameDocument3 pagesPyrolysis Polytetrafluoroethylene: A SameIsadora Berlanga MoraNo ratings yet
- Exploring The Limits of Branching and Gel Content in The Emulsion (2006)Document6 pagesExploring The Limits of Branching and Gel Content in The Emulsion (2006)iysernaNo ratings yet
- Os Coll. Vol. 6 P175-PtabDocument5 pagesOs Coll. Vol. 6 P175-Ptabsunil_vaman_joshiNo ratings yet
- Chem of Hetero Comp - MajorDocument4 pagesChem of Hetero Comp - MajorUnique AcademyNo ratings yet
- A-Oxygenated Crotyltitanium and Dyotropic Rearrangement in The Total Synthesis of DiscodermolideDocument5 pagesA-Oxygenated Crotyltitanium and Dyotropic Rearrangement in The Total Synthesis of Discodermolidefrancescomaugeri1986No ratings yet
- Sdarticle 18 PDFDocument8 pagesSdarticle 18 PDFPpa Gpat AmitNo ratings yet
- Main ArticleIodine Catalyzed One Pot Synthesis of 4-Iodo-3-Phenylbenzo (B) (1,6) Naphthyridine From Alkynylquinoline-3-Carbaldehydes, Via Imino Iodization of Alkynes.Document4 pagesMain ArticleIodine Catalyzed One Pot Synthesis of 4-Iodo-3-Phenylbenzo (B) (1,6) Naphthyridine From Alkynylquinoline-3-Carbaldehydes, Via Imino Iodization of Alkynes.Ashish TewariNo ratings yet
- Morphine Total SynthesisDocument3 pagesMorphine Total SynthesisLeidy Tatiana Lievano RubioNo ratings yet
- Preparation of Arylbutadiyne Derivatives by Silyl Migration Inspired by The Mukaiyama ProtocolDocument4 pagesPreparation of Arylbutadiyne Derivatives by Silyl Migration Inspired by The Mukaiyama ProtocolRaul tejadaNo ratings yet
- Brimer 1998Document7 pagesBrimer 1998Artem KulikovNo ratings yet
- An Improved Asymmetric Synthetic Route To D-Biotin Via Hoffmann-Roche Lactone-Thiolactone ApproachDocument4 pagesAn Improved Asymmetric Synthetic Route To D-Biotin Via Hoffmann-Roche Lactone-Thiolactone ApproachPaulo HenriqueNo ratings yet
- Ring-Forming Polymerizations Pt B 2: Heterocyclic RingsFrom EverandRing-Forming Polymerizations Pt B 2: Heterocyclic RingsRating: 5 out of 5 stars5/5 (1)
- Superabsorbent SynthesisDocument6 pagesSuperabsorbent SynthesisNikol TorresNo ratings yet
- Yokozawa 2000Document10 pagesYokozawa 2000h.sinner671No ratings yet
- Investigation of The False Positive Results of Dithiocarbamate Pesticides Due To Endogenous Nonpathogenic Carbon Disulfide in Organically Grown Plants (Moringa Oleifera)Document7 pagesInvestigation of The False Positive Results of Dithiocarbamate Pesticides Due To Endogenous Nonpathogenic Carbon Disulfide in Organically Grown Plants (Moringa Oleifera)Sabrina JonesNo ratings yet
- HGFDEDocument4 pagesHGFDEWalid Ebid ElgammalNo ratings yet
- Kamimura Et Al-2008-Polymers For Advanced TechnologiesDocument5 pagesKamimura Et Al-2008-Polymers For Advanced TechnologiesMonique PadovanNo ratings yet
- 1 PBDocument6 pages1 PBhatet77570No ratings yet
- 1999 Prochazkova Determinación Cuantitiva de Quitosano Por NinhidrinaDocument8 pages1999 Prochazkova Determinación Cuantitiva de Quitosano Por NinhidrinaRodrigo MiguelNo ratings yet
- Grignard ReactionDocument6 pagesGrignard Reactionjayhov86% (7)
- 3959 FTPDocument9 pages3959 FTPCabNo ratings yet
- J Tet 2007 11 076Document12 pagesJ Tet 2007 11 076AayuNo ratings yet
- Jurnal Sintesis Senyawa ObatDocument8 pagesJurnal Sintesis Senyawa ObatRizqy MannoNo ratings yet
- Improved Version of The Fischer-Zach Synthesis of Glycals: Vitamin B-12 Catalyzed Reductive Elimination of Glycosyl BromidesDocument2 pagesImproved Version of The Fischer-Zach Synthesis of Glycals: Vitamin B-12 Catalyzed Reductive Elimination of Glycosyl BromidesscadvijayNo ratings yet
- Trimethylamine N-Oxide: Me NODocument4 pagesTrimethylamine N-Oxide: Me NOVeneta GizdakovaNo ratings yet
- Analysis of Coconut Oil-Diethanolamine Condensates by Gas ChromatographyDocument4 pagesAnalysis of Coconut Oil-Diethanolamine Condensates by Gas ChromatographyAle MacedoNo ratings yet
- Analysis of Coconut Oil-Diethanolamine Condensates by Gas Chromatography PDFDocument4 pagesAnalysis of Coconut Oil-Diethanolamine Condensates by Gas Chromatography PDFAle MacedoNo ratings yet
- Synthesis of Polyamides From P-Xylylene GlycolDocument6 pagesSynthesis of Polyamides From P-Xylylene GlycolVirender KumarNo ratings yet
- Tetrahedron Letters 2002, 58, 6351-6358Document8 pagesTetrahedron Letters 2002, 58, 6351-6358Ricardo HernandezNo ratings yet
- B-H Bond ActivationDocument8 pagesB-H Bond ActivationLuna RosieNo ratings yet
- Síntese e Polimerização de Brometo de N, N-Dialil MorfolínioDocument8 pagesSíntese e Polimerização de Brometo de N, N-Dialil MorfolínioHeloisa Gabriele Miranda da SilvaNo ratings yet
- Improving The Sustainability of Biodiesel ProductionDocument1 pageImproving The Sustainability of Biodiesel ProductionVanessa Rios NolayaNo ratings yet
- Microwave SynthesisDocument4 pagesMicrowave SynthesisFatema HussainNo ratings yet
- Production of Biosurfactant From Torulopsis: BombicolaDocument4 pagesProduction of Biosurfactant From Torulopsis: BombicolaSui GenerisNo ratings yet
- RChE PCP DIAG 4Document5 pagesRChE PCP DIAG 4Paulo Emmanuele BetitaNo ratings yet
- Atom Transfer Radical Polymerization of Acrylated Epoxidized Soybean OilDocument1 pageAtom Transfer Radical Polymerization of Acrylated Epoxidized Soybean OilSandra AristizábalNo ratings yet
- Control of Reactivity of Novolac Resins: The Use of 3,4-Dihydro-2h-Pyran As A Labile Protecting GroupDocument7 pagesControl of Reactivity of Novolac Resins: The Use of 3,4-Dihydro-2h-Pyran As A Labile Protecting GroupQuế NghiNo ratings yet
- A New Reagent To Access Methyl SulfonesDocument10 pagesA New Reagent To Access Methyl Sulfonessaravana kumar.cNo ratings yet
- Prosedur Analisis CMCDocument4 pagesProsedur Analisis CMCardiantiresiNo ratings yet
- Total Synthesis of Pederin, A Potent Insect Toxin: The Ef®cient Synthesis of The Right Half, (1) - BenzoylpedamideDocument7 pagesTotal Synthesis of Pederin, A Potent Insect Toxin: The Ef®cient Synthesis of The Right Half, (1) - BenzoylpedamideHY-11 Đỗ Quốc TiệpNo ratings yet
- Percent Yield: Chemfile Mini-Guide To Problem SolvingDocument11 pagesPercent Yield: Chemfile Mini-Guide To Problem SolvingdhavaleshNo ratings yet
- Liu Et AlDocument9 pagesLiu Et AlSylvie jNo ratings yet
- Treatment of Sleep Disturbances in Post-Traumatic Stress Disorder: A Review of The LiteratureDocument10 pagesTreatment of Sleep Disturbances in Post-Traumatic Stress Disorder: A Review of The LiteratureSylvie jNo ratings yet
- Is Brain-Derived Neurotrophic Factor (BDNF) Val66Met Polymorphism Associated With Obsessive-Compulsive Disorder? A Meta-AnalysisDocument7 pagesIs Brain-Derived Neurotrophic Factor (BDNF) Val66Met Polymorphism Associated With Obsessive-Compulsive Disorder? A Meta-AnalysisSylvie jNo ratings yet
- Genetic and Environmental in Uences On Obsessive-Compulsive DisorderDocument11 pagesGenetic and Environmental in Uences On Obsessive-Compulsive DisorderSylvie jNo ratings yet
- The Benefits of Brassica Vegetables On Human HealthDocument13 pagesThe Benefits of Brassica Vegetables On Human HealthMikhaela Czharene FormosoNo ratings yet
- Sundow Polymers Co., LTD.: Surpren SN232Document2 pagesSundow Polymers Co., LTD.: Surpren SN232Phuong The NguyenNo ratings yet
- SCH 511-Biosynthesis of Terpenoids-IntroductionDocument30 pagesSCH 511-Biosynthesis of Terpenoids-IntroductionЕгяна КазимомаNo ratings yet
- Pesticides-Fertilizers Interaction On SoilDocument21 pagesPesticides-Fertilizers Interaction On SoilDreamleague SoccorNo ratings yet
- Life Sciences Grade 12 Term 1 Week 1 - 2021Document4 pagesLife Sciences Grade 12 Term 1 Week 1 - 2021Lucky Jr MonobeNo ratings yet
- Ecocert Detergents (Home Care)Document6 pagesEcocert Detergents (Home Care)Debra McCollamNo ratings yet
- Liver Function Tests... ROHITDocument61 pagesLiver Function Tests... ROHITJr Jc Rohit SahNo ratings yet
- Fundamentals of Rganic Chemistry Note For Year I PharmacyDocument151 pagesFundamentals of Rganic Chemistry Note For Year I PharmacySelam 1No ratings yet
- Synthetics, Mineral Oils, and Bio-Based Lubricants Chemistry and Technology, Third EditionDocument33 pagesSynthetics, Mineral Oils, and Bio-Based Lubricants Chemistry and Technology, Third EditionAceites MineralesNo ratings yet
- The Formaldehyde Method For DeterminingDocument2 pagesThe Formaldehyde Method For Determiningfahim khattakNo ratings yet
- 0 0 1211812181211ManufacturingProcessDocument17 pages0 0 1211812181211ManufacturingProcessLUISNo ratings yet
- 189922-Article Text-422623-1-10-20191227Document8 pages189922-Article Text-422623-1-10-20191227MohsenNo ratings yet
- Catalogo Consumibles Cromatografia de Gases y Espectrometria de Gases Parte3 PDFDocument218 pagesCatalogo Consumibles Cromatografia de Gases y Espectrometria de Gases Parte3 PDFTaranom SpNo ratings yet
- Prasanna Sajee Mock Test-Term 1 - Class 12Document8 pagesPrasanna Sajee Mock Test-Term 1 - Class 12Oreena DsouzaNo ratings yet
- Dow Science Paper-A Commercially Viable Solution Process To Control Long-Chain Branching in PolyethyleneDocument1 pageDow Science Paper-A Commercially Viable Solution Process To Control Long-Chain Branching in Polyethylenefengshaohua_gmailNo ratings yet
- Bioenergetics HandoutDocument1 pageBioenergetics HandoutRhaven GonzalesNo ratings yet
- Injera (An Ethnic, Traditional Staple Food of Ethiopia) : A Review On Traditional Practice To Scientific DevelopmentsDocument15 pagesInjera (An Ethnic, Traditional Staple Food of Ethiopia) : A Review On Traditional Practice To Scientific DevelopmentsTerikuwa MimoNo ratings yet
- Nomenclature of Chemical Compounds: Inorganic ChemistryDocument1 pageNomenclature of Chemical Compounds: Inorganic ChemistrybasimNo ratings yet
- CHEMISTRYQuestionDocument18 pagesCHEMISTRYQuestionGandhimathi mathiNo ratings yet
- Kulkarni Et Al. - 2022 - Role of Probiotics in Ruminant Nutrition As NaturaDocument15 pagesKulkarni Et Al. - 2022 - Role of Probiotics in Ruminant Nutrition As NaturaMATHILDE MAGRONo ratings yet
- Alternative Stain Remover Chapter 1Document10 pagesAlternative Stain Remover Chapter 1Jairelle NavaltaNo ratings yet
- Chimestry ProjectDocument28 pagesChimestry Projectpsyxs4tsv9No ratings yet
- Water-Borne polyurethane-Covestro-Bayhydrol A 2846Document27 pagesWater-Borne polyurethane-Covestro-Bayhydrol A 2846ichsan hakimNo ratings yet
- Study of UV Curing in The Wood Industry: Haider Osama Al-Mahdi My0001415Document8 pagesStudy of UV Curing in The Wood Industry: Haider Osama Al-Mahdi My0001415Adesh GurjarNo ratings yet
- TDS SHELL CATENEX SNR ARIZONAmotorsDocument2 pagesTDS SHELL CATENEX SNR ARIZONAmotorsİbrahim MutafogluNo ratings yet
- Applications of Metabolomics To Study Cancer MetabolismDocument13 pagesApplications of Metabolomics To Study Cancer Metabolismyylf2016No ratings yet
- Comparison of The Concentration of Phenolic Constituents SugarcaneDocument7 pagesComparison of The Concentration of Phenolic Constituents SugarcaneMolly0630No ratings yet