You might also like
- Impact of Invasive Sargassum on Native Macroalgal AssemblageDocument8 pagesImpact of Invasive Sargassum on Native Macroalgal AssemblageAlex Luna LlantirhuayNo ratings yet
- 10 1890@02-0431 PDFDocument8 pages10 1890@02-0431 PDFdindaNo ratings yet
- 2014 ReferenceDocument9 pages2014 ReferenceRayne LoefflerNo ratings yet
- A Reciprocal Inter-Habitat Transplant Reveals Changes in The Assemblage Structure of Macroinvertebrates Associated With The Sponge Halichondria Melanadocia. Avila y Briceño-Vera. 2018Document13 pagesA Reciprocal Inter-Habitat Transplant Reveals Changes in The Assemblage Structure of Macroinvertebrates Associated With The Sponge Halichondria Melanadocia. Avila y Briceño-Vera. 2018antony briceñoNo ratings yet
- Latitudinal Gradients SAX2001Document12 pagesLatitudinal Gradients SAX2001Jaypee MontonNo ratings yet
- Long-Term Changes in The Structure of Intertidal Assemblages After Invasion by Sargassum Muticum (Phaeophyta)Document8 pagesLong-Term Changes in The Structure of Intertidal Assemblages After Invasion by Sargassum Muticum (Phaeophyta)Alex Luna LlantirhuayNo ratings yet
- 37Document17 pages37arindaaa432No ratings yet
- Adaptive Bleaching Hypothesis Experimental TestsDocument8 pagesAdaptive Bleaching Hypothesis Experimental TestsChuk DiogoNo ratings yet
- Ecology of Free Living Marine NematodesDocument44 pagesEcology of Free Living Marine Nematodescitlali_DANo ratings yet
- Journal of Arid Environments: J.L. Mora, R. LázaroDocument10 pagesJournal of Arid Environments: J.L. Mora, R. LázaroEder ChaveiroNo ratings yet
- 2021 Global Gradients in Intertidal SpeciesDocument17 pages2021 Global Gradients in Intertidal SpeciesMauricio OrósticaNo ratings yet
- Articulo4 Patrones Biodiversidad PecesDocument7 pagesArticulo4 Patrones Biodiversidad PecesJulian RodriguezNo ratings yet
- Birkhofer Henschel Lubin 2012 SeothyrasurveyDocument7 pagesBirkhofer Henschel Lubin 2012 SeothyrasurveyAhmad Aulia Bahrun AmieqNo ratings yet
- Fulltext Retno PakeDocument14 pagesFulltext Retno PakeBenu SutartoNo ratings yet
- 86 Marga Biol Invasions 2016Document16 pages86 Marga Biol Invasions 2016Rui Miguel CarvalhoNo ratings yet
- Gallé2017 Article ClimateChangeImpoverishesAndHoDocument9 pagesGallé2017 Article ClimateChangeImpoverishesAndHoElmo KochNo ratings yet
- 13 Esponjas Porifera Parque Nacional San Esteban 2010Document14 pages13 Esponjas Porifera Parque Nacional San Esteban 2010carmenNo ratings yet
- Long-Term Shifts in The Communities of Odonata Effect of Chance or Climate ChangeDocument6 pagesLong-Term Shifts in The Communities of Odonata Effect of Chance or Climate Changegabriel vila verde nunesNo ratings yet
- Deep-Sea Research Part IIDocument20 pagesDeep-Sea Research Part IINurul Matinni AbdullahNo ratings yet
- Fast Climatic Changes Place An Endemic Canary IslaDocument16 pagesFast Climatic Changes Place An Endemic Canary IslaIH QureshiNo ratings yet
- 2020 ReferenceDocument12 pages2020 ReferenceRayne LoefflerNo ratings yet
- Ecosystem Changes Associated With Grazing Intensity On The Punta Ninfas Rangelands of Patagonia, ArgentinaDocument6 pagesEcosystem Changes Associated With Grazing Intensity On The Punta Ninfas Rangelands of Patagonia, ArgentinaPaula BeNo ratings yet
- Airoldi - 1998 - Roles of Disturbance, Sediment Stress, and Substratum Retention On Spatial Dominance in Algal TurfDocument12 pagesAiroldi - 1998 - Roles of Disturbance, Sediment Stress, and Substratum Retention On Spatial Dominance in Algal Turfepidamus1980No ratings yet
- Positive and Negative Effects of A Threatened Parrotfish On Reef EcosystemsDocument10 pagesPositive and Negative Effects of A Threatened Parrotfish On Reef EcosystemsHaliliNo ratings yet
- Environmental Influences On Regional DeeDocument43 pagesEnvironmental Influences On Regional Deecitlali_DANo ratings yet
- Okey 2003Document26 pagesOkey 2003paulinatapia09uvNo ratings yet
- Marine Mammal-GaliciaDocument23 pagesMarine Mammal-GaliciaBeatrizCamposNo ratings yet
- MANTEL VERLO Palmaer Et AlDocument8 pagesMANTEL VERLO Palmaer Et AljorgepierNo ratings yet
- Mechanisms of succession along the emersion gradient in intertidal rocky shore assemblagesDocument14 pagesMechanisms of succession along the emersion gradient in intertidal rocky shore assemblagesAlex Luna LlantirhuayNo ratings yet
- Diversity of Soil Fauna and Ecosystem FunctionDocument14 pagesDiversity of Soil Fauna and Ecosystem Functiontariqulmasud12No ratings yet
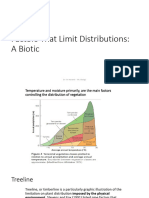
- Week - 5. Factors That Limit Distributions.a Biotic. NicheDocument31 pagesWeek - 5. Factors That Limit Distributions.a Biotic. NicheAnnisa VeterinerianNo ratings yet
- s10021 008 9191 7Document15 pagess10021 008 9191 7jvemgenhariaNo ratings yet
- Animal Habitat Quality and Ecosystem Functioning: Exploring Seasonal Patterns Using NdviDocument17 pagesAnimal Habitat Quality and Ecosystem Functioning: Exploring Seasonal Patterns Using NdvimiramontesNo ratings yet
- Kandelia Candel (L.) Druce in The PhilippinesDocument12 pagesKandelia Candel (L.) Druce in The Philippinestikz1567% (3)
- Sewage Impact on Polychaete Composition in Mussel BedsDocument10 pagesSewage Impact on Polychaete Composition in Mussel BedspablofedericciNo ratings yet
- Structure and Diversity of Intertidal Benthic Diatom Assemblages in Contrasting Shores: A Case Study From The Tagus EstuaryDocument13 pagesStructure and Diversity of Intertidal Benthic Diatom Assemblages in Contrasting Shores: A Case Study From The Tagus EstuaryCris FischerNo ratings yet
- Biodiversity Loss and Ecosystem Functioning: Distinguishing Between Number and Identity of SpeciesDocument14 pagesBiodiversity Loss and Ecosystem Functioning: Distinguishing Between Number and Identity of SpeciesLisa HanifahNo ratings yet
- Asquith e Chang 2005 - DSDocument12 pagesAsquith e Chang 2005 - DSBeatrizNo ratings yet
- Ecological Impacts of The Austral-Most Population of Crassostrea Gigas in South America: A Matter of Time?Document9 pagesEcological Impacts of The Austral-Most Population of Crassostrea Gigas in South America: A Matter of Time?Sarita EmmanuelNo ratings yet
- Full TextDocument12 pagesFull TextNoor HayatNo ratings yet
- Butterflies and Vegetation in Restored Gullies of Different Ages at The Colombian Western AndesDocument18 pagesButterflies and Vegetation in Restored Gullies of Different Ages at The Colombian Western Andeswretch706No ratings yet
- 11.1.124-Rojas-Soto et al., 2021Document10 pages11.1.124-Rojas-Soto et al., 2021Diego BaldoNo ratings yet
- Evegetación de Áreas Degradadas Con Especies Nativas: Antonio Daniel DalmassoDocument23 pagesEvegetación de Áreas Degradadas Con Especies Nativas: Antonio Daniel DalmassoAna Laura NavasNo ratings yet
- Walkup Et Al. (2017)Document15 pagesWalkup Et Al. (2017)Ruben FernandezNo ratings yet
- 11-35 - 2 Trama (175-89)Document15 pages11-35 - 2 Trama (175-89)flortramaNo ratings yet
- Urbina-Cardona Et Al 2006 BiolconsDocument15 pagesUrbina-Cardona Et Al 2006 BiolconsDaniel VásquezNo ratings yet
- 2 Tiselius and Moller JPR 2017 Community Cascades Caused by CtenophoreDocument9 pages2 Tiselius and Moller JPR 2017 Community Cascades Caused by CtenophoreRubyHudkinsNo ratings yet
- Thermal Tolerance of Mediterranean Marine MacrophyDocument12 pagesThermal Tolerance of Mediterranean Marine MacrophyAfonso PrestesNo ratings yet
- Mineur HALDocument19 pagesMineur HALPaola Andrea Franco MunozNo ratings yet
- Journal of Experimental Marine Biology and Ecology: Darlan J.B. Simith, Karen DieleDocument7 pagesJournal of Experimental Marine Biology and Ecology: Darlan J.B. Simith, Karen DieleharisankarhsNo ratings yet
- Macroalgal Species Diversity and Biomass of Subtidal Communities of São MiguelDocument11 pagesMacroalgal Species Diversity and Biomass of Subtidal Communities of São MiguelGuillermo Menéndez GarcíaNo ratings yet
- Easton Et Al. 2018. Effects of Herbivores, Wave Exposure and Depth On Benthic Coral Communities of The Easter Island EcoregionDocument10 pagesEaston Et Al. 2018. Effects of Herbivores, Wave Exposure and Depth On Benthic Coral Communities of The Easter Island Ecoregionhugo zeballosNo ratings yet
- TMP 4 B9 ADocument99 pagesTMP 4 B9 AFrontiersNo ratings yet
- Connell Etal1997Document28 pagesConnell Etal1997Princy LivingstonNo ratings yet
- Importancia Funcional de Comunidades Bentonicas Sanchis2021Document13 pagesImportancia Funcional de Comunidades Bentonicas Sanchis2021j.salazar.somNo ratings yet
- Jellyfish hotspots may explain 22.5% of leatherback turtle distributionDocument6 pagesJellyfish hotspots may explain 22.5% of leatherback turtle distributionBengt HörbergNo ratings yet
- Cox Et Al. 2019 Infaunal Community Responses To Ancient Clam GardensDocument12 pagesCox Et Al. 2019 Infaunal Community Responses To Ancient Clam GardensTrade TechnologiesNo ratings yet
- The Role of Highly Mobile Crab Predators in The Intertidal Zonation of Their Gastropod PreyDocument25 pagesThe Role of Highly Mobile Crab Predators in The Intertidal Zonation of Their Gastropod PreyPrincy CaesarNo ratings yet
- Limnetica 36 1 P 87Document12 pagesLimnetica 36 1 P 87Manuel PérezNo ratings yet
- Silver and Polymetallic Projects in Prolific Historic Mining DistrictsDocument23 pagesSilver and Polymetallic Projects in Prolific Historic Mining DistrictsAlex Luna LlantirhuayNo ratings yet
- Mechanisms of succession along the emersion gradient in intertidal rocky shore assemblagesDocument14 pagesMechanisms of succession along the emersion gradient in intertidal rocky shore assemblagesAlex Luna LlantirhuayNo ratings yet
- Size Structure and Dynamics in A Population of (Phaeophyceae)Document9 pagesSize Structure and Dynamics in A Population of (Phaeophyceae)Alex Luna LlantirhuayNo ratings yet
- Long-Term Changes in The Structure of Intertidal Assemblages After Invasion by Sargassum Muticum (Phaeophyta)Document8 pagesLong-Term Changes in The Structure of Intertidal Assemblages After Invasion by Sargassum Muticum (Phaeophyta)Alex Luna LlantirhuayNo ratings yet
- Baldosas PDFDocument10 pagesBaldosas PDFAlex Luna LlantirhuayNo ratings yet
- Resource Availability and Invasibility in An Intertidal Macroalgal AssemblageDocument10 pagesResource Availability and Invasibility in An Intertidal Macroalgal AssemblageAlex Luna LlantirhuayNo ratings yet
- Three Days To SeeDocument2 pagesThree Days To SeeMae MejaresNo ratings yet
- Chem 6AL Syllabus Winter 2021Document5 pagesChem 6AL Syllabus Winter 2021John SmithNo ratings yet
- Survey of Accounting 6th Edition Warren Solutions ManualDocument17 pagesSurvey of Accounting 6th Edition Warren Solutions Manualdevinsmithddsfzmioybeqr100% (19)
- Evidence OutlineDocument2 pagesEvidence OutlineMalia JordanNo ratings yet
- Playfair Cipher: Cipher Is A Manual Symmetric Encryption Technique and Was The First LiteralDocument9 pagesPlayfair Cipher: Cipher Is A Manual Symmetric Encryption Technique and Was The First LiteralJOHN CHARLASNo ratings yet
- Comparative ParadigmDocument4 pagesComparative ParadigmJovi Floresca AberinNo ratings yet
- Language Learning Enhanced by Music and SongDocument7 pagesLanguage Learning Enhanced by Music and SongNina Hudson100% (2)
- F K Up This Book Sample PDFDocument38 pagesF K Up This Book Sample PDFBlack & White PublishingNo ratings yet
- Micro To MACRO Mathematical Modelling in Soil Mechanics (Trends in Mathematics)Document407 pagesMicro To MACRO Mathematical Modelling in Soil Mechanics (Trends in Mathematics)Renova HnNo ratings yet
- Sheet-Pan Salmon and Broccoli With Sesame and Ginger: by Lidey HeuckDocument2 pagesSheet-Pan Salmon and Broccoli With Sesame and Ginger: by Lidey HeuckllawNo ratings yet
- Conciliar Post Reference GuideDocument9 pagesConciliar Post Reference GuideBenjamin WinterNo ratings yet
- The LOMA Weekly Herald Volume 1 Issue 6Document6 pagesThe LOMA Weekly Herald Volume 1 Issue 6LOMA MaksNo ratings yet
- 515 AMA Must DO QuestionsDocument217 pages515 AMA Must DO QuestionsVenkataRajuNo ratings yet
- Experiment No. 2. Frequency Shift Keying (FSK) and Digital CommunicationsDocument15 pagesExperiment No. 2. Frequency Shift Keying (FSK) and Digital CommunicationsAmir Mahmood RehmaniNo ratings yet
- Letter To Editor NDocument5 pagesLetter To Editor NNavya AgarwalNo ratings yet
- Finaflex Main Catalog 2022Document50 pagesFinaflex Main Catalog 2022Benlee Calderón LimaNo ratings yet
- Exercise For FitnessDocument44 pagesExercise For FitnessSheena Mae Sube PoNo ratings yet
- KZN OUTER WEST Development ProjectsDocument25 pagesKZN OUTER WEST Development ProjectsDurban Chamber of Commerce and IndustryNo ratings yet
- 9331 DCCBDocument5 pages9331 DCCBgopimodalavalasaNo ratings yet
- Food Safety Culture Webinar SLIDESDocument46 pagesFood Safety Culture Webinar SLIDESAto Kwamena PaintsilNo ratings yet
- (Vampire Library) Gail B. Stewart - Vampires. Do They Exist - ReferencePoint Press (2011)Document80 pages(Vampire Library) Gail B. Stewart - Vampires. Do They Exist - ReferencePoint Press (2011)Jerry KanneNo ratings yet
- Moot Problem For ICA - NMIMS - June 2021Document3 pagesMoot Problem For ICA - NMIMS - June 2021AayushiNo ratings yet
- REVISED 2023-2025 LIPH Appraisal Checklist Rating Scale v5 9.12Document11 pagesREVISED 2023-2025 LIPH Appraisal Checklist Rating Scale v5 9.12Joshua AssinNo ratings yet
- Part of Speech QuestionsDocument4 pagesPart of Speech QuestionsNi Luh Ari Kusumawati0% (1)
- Magh Bihu or Maghar DomahiDocument8 pagesMagh Bihu or Maghar Domahihackdarenot4No ratings yet
- EthicsDocument10 pagesEthicsEssi Chan100% (4)
- Awards and Honors 2018Document79 pagesAwards and Honors 2018rajinder345No ratings yet
- AISD Fees Structure for 2023-24 Academic YearDocument2 pagesAISD Fees Structure for 2023-24 Academic YearKawsar AlamNo ratings yet
- Integrated BMSDocument14 pagesIntegrated BMSjim.walton100% (5)
- Grade 9: International Junior Math OlympiadDocument13 pagesGrade 9: International Junior Math OlympiadLong Văn Trần100% (1)
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseFrom EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseRating: 3.5 out of 5 stars3.5/5 (69)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessFrom EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo ratings yet
- World of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsFrom EverandWorld of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsRating: 4 out of 5 stars4/5 (222)
- The Other End of the Leash: Why We Do What We Do Around DogsFrom EverandThe Other End of the Leash: Why We Do What We Do Around DogsRating: 5 out of 5 stars5/5 (63)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorFrom EverandThe Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorRating: 4.5 out of 5 stars4.5/5 (137)
- Come Back, Como: Winning the Heart of a Reluctant DogFrom EverandCome Back, Como: Winning the Heart of a Reluctant DogRating: 3.5 out of 5 stars3.5/5 (10)
- Why Fish Don't Exist: A Story of Loss, Love, and the Hidden Order of LifeFrom EverandWhy Fish Don't Exist: A Story of Loss, Love, and the Hidden Order of LifeRating: 4.5 out of 5 stars4.5/5 (699)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- When You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsFrom EverandWhen You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsRating: 3.5 out of 5 stars3.5/5 (13)
- Fire Season: Field Notes from a Wilderness LookoutFrom EverandFire Season: Field Notes from a Wilderness LookoutRating: 4 out of 5 stars4/5 (142)
- The Big, Bad Book of Botany: The World's Most Fascinating FloraFrom EverandThe Big, Bad Book of Botany: The World's Most Fascinating FloraRating: 3 out of 5 stars3/5 (10)
- The Secret Life of Lobsters: How Fishermen and Scientists Are Unraveling the Mysteries of Our Favorite CrustaceanFrom EverandThe Secret Life of Lobsters: How Fishermen and Scientists Are Unraveling the Mysteries of Our Favorite CrustaceanNo ratings yet
- Gathering Moss: A Natural and Cultural History of MossesFrom EverandGathering Moss: A Natural and Cultural History of MossesRating: 4.5 out of 5 stars4.5/5 (347)
- The Mind of Plants: Narratives of Vegetal IntelligenceFrom EverandThe Mind of Plants: Narratives of Vegetal IntelligenceRating: 4.5 out of 5 stars4.5/5 (11)
- Last Child in the Woods: Saving Our Children From Nature-Deficit DisorderFrom EverandLast Child in the Woods: Saving Our Children From Nature-Deficit DisorderRating: 4 out of 5 stars4/5 (283)
- Spoiled Rotten America: Outrages of Everyday LifeFrom EverandSpoiled Rotten America: Outrages of Everyday LifeRating: 3 out of 5 stars3/5 (19)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (593)