You might also like
- bioenergetic NDocument3 pagesbioenergetic NReema Genice BayabordaNo ratings yet
- Module 5. Bacterial MetabolismDocument22 pagesModule 5. Bacterial MetabolismIRENE SEBASTIANNo ratings yet
- Laws of Thermodynamics As Related To BiologyDocument2 pagesLaws of Thermodynamics As Related To BiologyMohammed MohammedNo ratings yet
- Energy in BiologyDocument15 pagesEnergy in BiologyJay BetaizarNo ratings yet
- An Introduction To MetabolismDocument9 pagesAn Introduction To MetabolismsamNo ratings yet
- Cellular Energy Currency: ATP Powers MetabolismDocument29 pagesCellular Energy Currency: ATP Powers MetabolismClaire WeerepasNo ratings yet
- Chemistry of LifeDocument15 pagesChemistry of Lifeapi-233187566No ratings yet
- Bioenergetics KOMDocument4 pagesBioenergetics KOMGayathri potnuru JioNo ratings yet
- Thermodynamics and BiologyDocument4 pagesThermodynamics and BiologyAlice RiddleNo ratings yet
- Green Minimalist The Environtment PresentationDocument11 pagesGreen Minimalist The Environtment PresentationJimmy Q MadroneroNo ratings yet
- AP Biology Unit 3 Student NotesDocument46 pagesAP Biology Unit 3 Student NotesJ15No ratings yet
- Bioenergetics ExplainedDocument29 pagesBioenergetics ExplainedMjNo ratings yet
- How cells obtain energy from foodDocument4 pagesHow cells obtain energy from foodBuzoianu Maria100% (1)
- Big Idea 2 - ReviewDocument23 pagesBig Idea 2 - ReviewGeoffrey BraunNo ratings yet
- Energy - MetabolismDocument45 pagesEnergy - MetabolismAngel GustasianaNo ratings yet
- Energy Transfer in Cell During Exercise and Oxygen Metabolism and Transport - PalakDocument85 pagesEnergy Transfer in Cell During Exercise and Oxygen Metabolism and Transport - Palakpalooo4No ratings yet
- Biology: a QuickStudy Laminated Reference GuideFrom EverandBiology: a QuickStudy Laminated Reference GuideRating: 3 out of 5 stars3/5 (2)
- PLANT METABOLISMDocument15 pagesPLANT METABOLISMnegiineha2No ratings yet
- Hierarchy of Biolgical OrganizationDocument21 pagesHierarchy of Biolgical OrganizationSal Sabeela RahmanNo ratings yet
- CHAPTER 3 Cellular Energy Transformation v1Document70 pagesCHAPTER 3 Cellular Energy Transformation v1Dhanrence AmuraoNo ratings yet
- Lesson 1 - EcosystemDocument3 pagesLesson 1 - EcosystemPaul Assie RosarioNo ratings yet
- Topic: Bioenergetics: Living OrganismsDocument9 pagesTopic: Bioenergetics: Living OrganismsNaiomiNo ratings yet
- Biology Notes - Module 2 Part 2Document19 pagesBiology Notes - Module 2 Part 2belindaNo ratings yet
- An Introduction To Metabolism: Lecture OutlineDocument12 pagesAn Introduction To Metabolism: Lecture Outlinehaha_le12No ratings yet
- Sytem Concepts and InteractionDocument32 pagesSytem Concepts and InteractionRhealyn RobledoNo ratings yet
- Bioenergetics ExplainedDocument7 pagesBioenergetics ExplainedBrave MitraNo ratings yet
- Thermodynamics and Its ImplicationDocument5 pagesThermodynamics and Its ImplicationMadan ThapaNo ratings yet
- 1.introduction To MetabolismDocument7 pages1.introduction To MetabolismProtusha RakshitNo ratings yet
- Unit 3: Energy Transformation General Biology 1 2 Quarter 1. ATP-ADP CycleDocument10 pagesUnit 3: Energy Transformation General Biology 1 2 Quarter 1. ATP-ADP CycleSophia AbatayNo ratings yet
- Module 5Document8 pagesModule 5mirai desuNo ratings yet
- Lesson 1: Introduction To Bioenergetics: Review QuestionsDocument2 pagesLesson 1: Introduction To Bioenergetics: Review QuestionsBea SamonteNo ratings yet
- Energy Flow in An Ecosystem01Document14 pagesEnergy Flow in An Ecosystem01ALIGNDALNo ratings yet
- Botany Module 4.Document20 pagesBotany Module 4.minsumainlhsNo ratings yet
- Microbial ChemistryDocument10 pagesMicrobial ChemistryChineduNo ratings yet
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- Applicability of 1 and 2 Laws of Thermodynamics On Living CellsDocument3 pagesApplicability of 1 and 2 Laws of Thermodynamics On Living CellsMathiasNo ratings yet
- Biochemistry Lecture 1: Cell Structure and EnergyDocument62 pagesBiochemistry Lecture 1: Cell Structure and EnergyhideNo ratings yet
- Chapter 4 Energy and EnzymesDocument19 pagesChapter 4 Energy and EnzymesfatimaNo ratings yet
- Bioenergetics ExplainedDocument7 pagesBioenergetics ExplainedGemay DanglayNo ratings yet
- Written Report On MicrophyDocument13 pagesWritten Report On MicrophyAustin Dela CruzNo ratings yet
- Microbial Physiology and GeneticsDocument3 pagesMicrobial Physiology and GeneticsYna Joy B. LigatNo ratings yet
- Non-Living and Living Components Environ PDFDocument22 pagesNon-Living and Living Components Environ PDFmatrixxxx420No ratings yet
- Topic 1 Notes 1Document6 pagesTopic 1 Notes 1api-293573854No ratings yet
- Handout in Biochem U1Document6 pagesHandout in Biochem U1Jenny RoseNo ratings yet
- Environmental Systems & Societies Notes: Topic 1: Systems and ModelsDocument9 pagesEnvironmental Systems & Societies Notes: Topic 1: Systems and ModelsVishnu SharmaNo ratings yet
- Lesson 2 The EcosystemDocument4 pagesLesson 2 The EcosystemMARIROSE ANCHETANo ratings yet
- Bio102 - EnergyDocument54 pagesBio102 - EnergyAbdul Hamed QaderiNo ratings yet
- ESS - Topic 1 NotesDocument6 pagesESS - Topic 1 NotesNicoleNo ratings yet
- Chapter 7: Energy and Ecosystems: SummaryDocument3 pagesChapter 7: Energy and Ecosystems: SummaryMerimaNo ratings yet
- 2.2 Communities and EcosystemsDocument53 pages2.2 Communities and EcosystemsGuzelle CooperNo ratings yet
- Chapter 8 - An Introduction To Metabolism NotesDocument12 pagesChapter 8 - An Introduction To Metabolism NotesDavid ThaiNo ratings yet
- General Biology 101Document7 pagesGeneral Biology 101Czarina AsisNo ratings yet
- 8th Grade ScienceDocument5 pages8th Grade ScienceMcDonaldNo ratings yet
- Definition of EcosystemDocument3 pagesDefinition of EcosystemManjunathan Sekar A2B PositiveNo ratings yet
- BioenergeticsDocument4 pagesBioenergeticsMahnoor MansoorNo ratings yet
- Thermodynamics of Chemical Processes in Living SystemsDocument16 pagesThermodynamics of Chemical Processes in Living SystemsJOSUE ZAKAYO KILAKOINo ratings yet
- Ecosystem Processes and BiogeochemistryDocument9 pagesEcosystem Processes and BiogeochemistryMoriel PradoNo ratings yet
- AP Bio Chapter 8Document5 pagesAP Bio Chapter 8Brendan Egan100% (2)
- Devlabdrawings - Exercise 4Document8 pagesDevlabdrawings - Exercise 4Art Julius D. HallazgoNo ratings yet
- Exercise 6. Chick Development: Egg To 28 Hours of IncubationDocument8 pagesExercise 6. Chick Development: Egg To 28 Hours of IncubationArt Julius D. HallazgoNo ratings yet
- Exercise 3 - Embryology: Art Julius Hallazgo Developmental Biology Laboratory - YaDocument5 pagesExercise 3 - Embryology: Art Julius Hallazgo Developmental Biology Laboratory - YaArt Julius D. HallazgoNo ratings yet
- An Advanced and Highly Intelligent Species of LifeDocument1 pageAn Advanced and Highly Intelligent Species of LifeArt Julius D. HallazgoNo ratings yet
- DevlecfruitflyforumDocument1 pageDevlecfruitflyforumArt Julius D. HallazgoNo ratings yet
- Art Hallazgo Gram Staining Exercise: Usp SharingDocument1 pageArt Hallazgo Gram Staining Exercise: Usp SharingArt Julius D. HallazgoNo ratings yet
- Ecolec Guide QuestionsDocument26 pagesEcolec Guide QuestionsArt Julius D. Hallazgo100% (1)
- Identification of Unknown Plant SpecimensDocument29 pagesIdentification of Unknown Plant SpecimensArt Julius D. HallazgoNo ratings yet
- Art Hallazgo Developmental Biology YA: Exercise 7: Chick Development: 33-60 Hours EmbryosDocument7 pagesArt Hallazgo Developmental Biology YA: Exercise 7: Chick Development: 33-60 Hours EmbryosArt Julius D. HallazgoNo ratings yet
- Sea Otters (Enhydra Lutris) : A Keystone Species: Submitted byDocument33 pagesSea Otters (Enhydra Lutris) : A Keystone Species: Submitted byArt Julius D. HallazgoNo ratings yet
- Lab Ex 1 - The CellDocument16 pagesLab Ex 1 - The CellArt Julius D. HallazgoNo ratings yet
- An Advanced and Highly Intelligent Species of LifeDocument1 pageAn Advanced and Highly Intelligent Species of LifeArt Julius D. HallazgoNo ratings yet
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- The Difference Between Homeostasis and MetabolismDocument1 pageThe Difference Between Homeostasis and MetabolismArt Julius D. HallazgoNo ratings yet
- nCoV Orientation As of 2 Feb 2020Document23 pagesnCoV Orientation As of 2 Feb 2020Art Julius D. HallazgoNo ratings yet
- When Performing ChromatographyDocument1 pageWhen Performing ChromatographyArt Julius D. HallazgoNo ratings yet
- LipidsDocument71 pagesLipidsArt Julius D. HallazgoNo ratings yet
- Catalase Enzyme LabDocument3 pagesCatalase Enzyme LabBruce0% (1)
- Genlab Exp1Document3 pagesGenlab Exp1Art Julius D. HallazgoNo ratings yet
- Rna TypesDocument2 pagesRna TypesArt Julius D. HallazgoNo ratings yet
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- MethodsDocument4 pagesMethodsGianne AbcedeNo ratings yet
- Kenshuquiz AkutalvarezquinonezDocument6 pagesKenshuquiz AkutalvarezquinonezArt Julius D. HallazgoNo ratings yet
- Lipids LabdiscussionDocument16 pagesLipids LabdiscussionArt Julius D. HallazgoNo ratings yet
- Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency DeficiencyDocument2 pagesDeficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency Deficiency DeficiencyArt Julius D. HallazgoNo ratings yet
- Chem Lab - 1 AnswersDocument2 pagesChem Lab - 1 AnswersArt Julius D. HallazgoNo ratings yet
- Analysis of SalivaDocument5 pagesAnalysis of SalivaYap JackyNo ratings yet
- Eng 5Document1 pageEng 5Art Julius D. HallazgoNo ratings yet
- Exp 2 - Post Lab BiochemDocument5 pagesExp 2 - Post Lab BiochemArt Julius D. HallazgoNo ratings yet
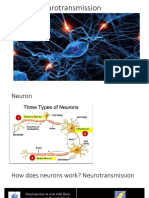
- Neurotransmission Processes and ReceptorsDocument66 pagesNeurotransmission Processes and ReceptorsSheik MohammedNo ratings yet
- Metabotropic Glutamate ReceptorsDocument9 pagesMetabotropic Glutamate Receptorslucia desantisNo ratings yet
- Chapter 4 Enzymes and VitaminsDocument9 pagesChapter 4 Enzymes and Vitaminsvictoria cablayNo ratings yet
- 1 EnzymesDocument30 pages1 EnzymesRohit pansareNo ratings yet
- Enzymology: Theresa May Chin TanDocument20 pagesEnzymology: Theresa May Chin TanVejerla PriyankaNo ratings yet
- Enzyme RegulationDocument1 pageEnzyme RegulationVea AngelesNo ratings yet
- 2021 Enzyme Biochemistry For Medical Students IIDocument146 pages2021 Enzyme Biochemistry For Medical Students IIJerome EkohNo ratings yet
- Biology Concepts and Connections Campbell 6th Edition Test BankDocument25 pagesBiology Concepts and Connections Campbell 6th Edition Test BankRichard RunnelsNo ratings yet
- Chapter 4 Biology Form 4: EnzymesDocument26 pagesChapter 4 Biology Form 4: EnzymesNurul Husna100% (1)
- Allosteric EnzymeDocument22 pagesAllosteric EnzymeAhmed ImranNo ratings yet
- BCH 201 DR Saliu Lecture NotesDocument19 pagesBCH 201 DR Saliu Lecture NotesAdekunle FestusNo ratings yet
- Week 2 - Enzyme Kinetics All - v2 - 3slide HandoutsDocument14 pagesWeek 2 - Enzyme Kinetics All - v2 - 3slide HandoutsFizza HussainNo ratings yet
- AnPhys3e Ch02 Test BankDocument22 pagesAnPhys3e Ch02 Test BankSuny Chavarria100% (2)
- WELCOME TO AP BIOLOGYDocument117 pagesWELCOME TO AP BIOLOGYRadha RamineniNo ratings yet
- Regulation of Cellular Respiration EnzymesDocument12 pagesRegulation of Cellular Respiration Enzymesdeepali_nih9585No ratings yet
- Enzyme Kinetics and InhibitionDocument25 pagesEnzyme Kinetics and InhibitionAstra BeckettNo ratings yet
- MCAT AAMC Content Outline - ScienceDocument165 pagesMCAT AAMC Content Outline - ScienceSumant SNo ratings yet
- Chapter 23Document53 pagesChapter 23haririNo ratings yet
- Module 4 biopharmaceutics overviewDocument8 pagesModule 4 biopharmaceutics overviewAzech Yam Ü100% (1)
- Chapter 8 Study Guide Ap BioDocument6 pagesChapter 8 Study Guide Ap BioIllan Chichportich100% (1)
- Biology ProjectDocument41 pagesBiology ProjectGanesan Siva67% (12)
- SBIA031 Study Guide 2024Document10 pagesSBIA031 Study Guide 2024Itumeleng SefaraNo ratings yet
- Enzymes PowerpointDocument39 pagesEnzymes PowerpointRizky Yudha IrawanNo ratings yet
- 2023-08-18 Enzymes KineticsDocument52 pages2023-08-18 Enzymes KineticsAjay MahalkaNo ratings yet
- Enzyme Inhibition/Enzyme InhibitorsDocument5 pagesEnzyme Inhibition/Enzyme InhibitorsFaria bukhariNo ratings yet
- ENVS 532: DR Assad Al-Thukair Associate ProfessorDocument20 pagesENVS 532: DR Assad Al-Thukair Associate ProfessorAbu Muhsin Al NgapakyNo ratings yet
- ENZYMES COMPLETE NOTES (UNIT 5 - B.Pharm 2nd Sem) PDFDocument15 pagesENZYMES COMPLETE NOTES (UNIT 5 - B.Pharm 2nd Sem) PDFBhavana Gangurde92% (36)
- BiochemistryDocument82 pagesBiochemistryAmritzz Paul83% (6)
- Enzymology NotesDocument5 pagesEnzymology NotesRyan Fortune AludaNo ratings yet