You might also like
- ART - Spectrophotometric Method For Determining GA in ..Document4 pagesART - Spectrophotometric Method For Determining GA in ..Azahara Linares100% (2)
- Kami Export - Omarion Fladger - Pedigree Genetics ProblemsDocument2 pagesKami Export - Omarion Fladger - Pedigree Genetics ProblemsOmarion FladgerNo ratings yet
- Duels and DuetsDocument253 pagesDuels and DuetsM Xenia0% (1)
- 10.12 - Horn - Combinatorial Selection in Amoebal Hosts Drives Evolution LegionellaDocument22 pages10.12 - Horn - Combinatorial Selection in Amoebal Hosts Drives Evolution LegionellamariangelestmNo ratings yet
- Quorum Sensing in African Trypanosomes: SciencedirectDocument6 pagesQuorum Sensing in African Trypanosomes: SciencedirectNithin SakthiNo ratings yet
- Chourasia Et Al 2020 Plasmodium Falciparum Clag9 Associated Pfrhoph Complex Is Involved in Merozoite Binding To HumanDocument14 pagesChourasia Et Al 2020 Plasmodium Falciparum Clag9 Associated Pfrhoph Complex Is Involved in Merozoite Binding To HumanArunaditya Deshmukh (Adi)No ratings yet
- Jir2018 6529681Document6 pagesJir2018 6529681Herry JapariNo ratings yet
- Hemoplasma Infection in Wild Captive CarnivoresDocument7 pagesHemoplasma Infection in Wild Captive CarnivoresEnfermagem com AlegriaNo ratings yet
- Virus Vectors IDocument14 pagesVirus Vectors IRatna Mutia KharismaNo ratings yet
- Lin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesDocument15 pagesLin, Z. (2018) - A Metalloprotease Homolog Venom Protein From A Parasitoid Wasp Suppresses The Toll Pathway in Host HemocytesJanina PuenteNo ratings yet
- Antigen Diversity in The Parasitic Bacterium AnaplasmaDocument9 pagesAntigen Diversity in The Parasitic Bacterium AnaplasmajmhechavarriaNo ratings yet
- Antimicrobial Peptides: University of Salerno Dept. of Pharmaceutical SciencesDocument7 pagesAntimicrobial Peptides: University of Salerno Dept. of Pharmaceutical Sciencesapi-3839948No ratings yet
- Vaccine Amazonensis8Document16 pagesVaccine Amazonensis8Eu Meu MesmoNo ratings yet
- Ijeb 47 (6) 412-423Document12 pagesIjeb 47 (6) 412-423Daniela MartinezNo ratings yet
- Infectious Diseases 2Document17 pagesInfectious Diseases 2JohnchubyNo ratings yet
- Host-Shifts Vs Cospeciation in Oomycete PathogensDocument21 pagesHost-Shifts Vs Cospeciation in Oomycete PathogensSergio D. PérezNo ratings yet
- Immune Escape Strategies MaleriaDocument7 pagesImmune Escape Strategies MaleriaAurelia WetondieNo ratings yet
- Sap RematikDocument7 pagesSap RematikOzada RasifaNo ratings yet
- Comparative Genomics and Molecular Analysis of Epidermophyton FloccosumDocument11 pagesComparative Genomics and Molecular Analysis of Epidermophyton Floccosumopus.mj1No ratings yet
- Camp Signalling in Trypanosomatids: Role in Pathogenesis and As A Drug TargetDocument7 pagesCamp Signalling in Trypanosomatids: Role in Pathogenesis and As A Drug TargetCamilo Ernesto Araujo BarabasNo ratings yet
- Breaking Down BiofilmsDocument3 pagesBreaking Down Biofilmsleila.hiloutNo ratings yet
- Immunopathogenesis of Pemphigus Vulgaris: A Brief ReviewDocument3 pagesImmunopathogenesis of Pemphigus Vulgaris: A Brief ReviewAnonymous SPrI0gbTr0No ratings yet
- Filaria PDFDocument6 pagesFilaria PDFanapangistiningsihNo ratings yet
- Anopheline Salivary Protein GenesDocument27 pagesAnopheline Salivary Protein Genesshweta pasiNo ratings yet
- Literature Review On Pseudomonas AeruginosaDocument5 pagesLiterature Review On Pseudomonas Aeruginosaamjatzukg100% (1)
- CANINE-Pemphigus Complex-Pathogenesis and Clinical PresentationDocument5 pagesCANINE-Pemphigus Complex-Pathogenesis and Clinical Presentationtaner_soysuren100% (4)
- Fimmu 11 00382Document19 pagesFimmu 11 00382Faiz SheeNo ratings yet
- 2021-Review-Phages To Modulate The Microbiome and ImmunityDocument16 pages2021-Review-Phages To Modulate The Microbiome and ImmunityCristian Felipe Sandoval QuiñonezNo ratings yet
- tmp85B6 TMPDocument7 pagestmp85B6 TMPFrontiersNo ratings yet
- 3 - Iabcr 20162 03 02 - 13 17Document6 pages3 - Iabcr 20162 03 02 - 13 17EMNo ratings yet
- Uropathogenic Proteus Mirabilis PDFDocument4 pagesUropathogenic Proteus Mirabilis PDFAttis PhrygiaNo ratings yet
- Malaria EnzimeDocument13 pagesMalaria EnzimeAnisahMahardianiNo ratings yet
- PDM Os 9Document15 pagesPDM Os 9Erdinawira RizkianiNo ratings yet
- Kima2007 PDFDocument10 pagesKima2007 PDFCLAUDIA QUINTERONo ratings yet
- Parasita No Intestino Do MosquitoDocument8 pagesParasita No Intestino Do MosquitoGustavo FelpeNo ratings yet
- 2016 Maintenance of Phenotypic Diversity Within A Set of Virulence Encoding Genes of The Malaria Parasite Plasmodium FalciparumDocument9 pages2016 Maintenance of Phenotypic Diversity Within A Set of Virulence Encoding Genes of The Malaria Parasite Plasmodium FalciparumSethawud ChaikitgosiyakulNo ratings yet
- International Journal For Parasitology: Musa A. Hassan, Aude-Anais Olijnik, Eva-Maria Frickel, Jeroen P. SaeijDocument8 pagesInternational Journal For Parasitology: Musa A. Hassan, Aude-Anais Olijnik, Eva-Maria Frickel, Jeroen P. SaeijwindaNo ratings yet
- Fantastic Yeasts and Where To Find Them: The Hidden Diversity of Dimorphic Fungal PathogensDocument9 pagesFantastic Yeasts and Where To Find Them: The Hidden Diversity of Dimorphic Fungal PathogensNithin SakthiNo ratings yet
- Fungos Entomopatogenico Interação Hospedeiro e VetorDocument58 pagesFungos Entomopatogenico Interação Hospedeiro e VetorGabriel Henrique Santos SilveiraNo ratings yet
- Pemhigus Lancet Schmidt2019Document13 pagesPemhigus Lancet Schmidt2019Faten HayderNo ratings yet
- Salmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008Document8 pagesSalmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008noordin MukasaNo ratings yet
- The Enigmatic PEPPE Multi-Gene Family of MycobacteDocument8 pagesThe Enigmatic PEPPE Multi-Gene Family of MycobacteNathan IbaleNo ratings yet
- Virulence Factors of Candida SpeciesDocument6 pagesVirulence Factors of Candida Specieslaura_ruiz_99No ratings yet
- Congenic Mice: Cutting Tools For Complex Immune Disorders: Ute C. Rogner and Philip AvnerDocument11 pagesCongenic Mice: Cutting Tools For Complex Immune Disorders: Ute C. Rogner and Philip AvnerJimit GandhiNo ratings yet
- Journal of MalariaDocument7 pagesJournal of MalariamyaNo ratings yet
- Sji 12533 PDFDocument7 pagesSji 12533 PDFanapangistiningsihNo ratings yet
- Accepted Manuscript: Molecular & Biochemical ParasitologyDocument25 pagesAccepted Manuscript: Molecular & Biochemical ParasitologyColo VoltaNo ratings yet
- Malaria: Jonathan Kidd Jennifer Koehl Heather Louch Edwin Wong Penelope WorthingtonDocument10 pagesMalaria: Jonathan Kidd Jennifer Koehl Heather Louch Edwin Wong Penelope WorthingtonraghvendrmNo ratings yet
- Patogenia de La ToxoplasmosisDocument10 pagesPatogenia de La ToxoplasmosisSasha de la CruzNo ratings yet
- Legionella PneumophilaDocument5 pagesLegionella PneumophilaOjambo FlaviaNo ratings yet
- Kuliah Penunjang Blok Imunologi: Paramasari Dirgahayu, DR., PHDDocument69 pagesKuliah Penunjang Blok Imunologi: Paramasari Dirgahayu, DR., PHDsekar tyasNo ratings yet
- Interactions of Fungal Pathogens With Phagocytes: Lars P. Erwig and Neil A. R. GowDocument14 pagesInteractions of Fungal Pathogens With Phagocytes: Lars P. Erwig and Neil A. R. GowPatrícia KellenNo ratings yet
- Pathogenesis of Fungal InfectionsDocument70 pagesPathogenesis of Fungal Infectionsማላያላም ማላያላም100% (15)
- Journal of Virology-2017-Reis-e02338-16.fullDocument6 pagesJournal of Virology-2017-Reis-e02338-16.fullSherly LudjiNo ratings yet
- Recent Buzz in Malaria Research: The FEBS Journal, Editorial Office, Cambridge, UKDocument4 pagesRecent Buzz in Malaria Research: The FEBS Journal, Editorial Office, Cambridge, UKVerlyanita SeptiariniNo ratings yet
- Infections of M. PneumoniaeDocument18 pagesInfections of M. PneumoniaeAndrés Nicolás Beltrán SeguraNo ratings yet
- Pathogens 11 00811Document5 pagesPathogens 11 00811octav drutaNo ratings yet
- 2004 Macrophages, Pathology and Parasite Persistence in Experimental Visceral LeishmaniasisDocument7 pages2004 Macrophages, Pathology and Parasite Persistence in Experimental Visceral LeishmaniasismclimacoNo ratings yet
- 570 1280 1 SMDocument10 pages570 1280 1 SMRachmat Budi PrasetyoNo ratings yet
- BOHOL Plasmodium MalariaeDocument10 pagesBOHOL Plasmodium MalariaeMabz BoholNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- Management of Infections in the Immunocompromised HostFrom EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalNo ratings yet
- A Micro Biuret Method F O R Protein DeterminationDocument5 pagesA Micro Biuret Method F O R Protein DeterminationNjani DsiNo ratings yet
- Kurva AmamiDocument2 pagesKurva AmamiNjani DsiNo ratings yet
- AbsenDocument1 pageAbsenNjani DsiNo ratings yet
- Taqman Real-Time PCR For Detecting Bovine Viral Diarrhea VirusDocument9 pagesTaqman Real-Time PCR For Detecting Bovine Viral Diarrhea VirusNjani DsiNo ratings yet
- 3 - Plasma Membrane and Transport MechanismsDocument67 pages3 - Plasma Membrane and Transport MechanismsThom PaglinawanNo ratings yet
- 300 One Word Substitutions Asked in SSC IBPS UPSC ExamDocument12 pages300 One Word Substitutions Asked in SSC IBPS UPSC ExamRashid AliNo ratings yet
- Ch8 (C)Document28 pagesCh8 (C)fkjujNo ratings yet
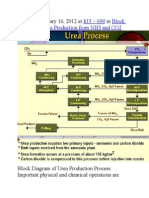
- Published January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2Document9 pagesPublished January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2himanshuchawla654No ratings yet
- 〈89.1〉 Collagenase IDocument5 pages〈89.1〉 Collagenase IAvijit HazraNo ratings yet
- Temperament ThesisDocument4 pagesTemperament ThesisBuyCheapPaperOnlineSingapore100% (2)
- Characterization of Tannia PlantDocument104 pagesCharacterization of Tannia PlantSolomon FantawNo ratings yet
- Gender and PsychotherapyDocument12 pagesGender and PsychotherapyEhh EnNaNo ratings yet
- Weekly Home Learning Plan: Day/Time Learning Competency Learning Tasks Mode of DeliveryDocument4 pagesWeekly Home Learning Plan: Day/Time Learning Competency Learning Tasks Mode of DeliveryEricha SolomonNo ratings yet
- Grade 5, Science CurriculumDocument3 pagesGrade 5, Science CurriculumJoel C. Yuvienco67% (3)
- UltraSonic Processors - CatalogDocument2 pagesUltraSonic Processors - Catalogdéborah_rosalesNo ratings yet
- RPT SC Year 2 (DLP) 2022-2023Document21 pagesRPT SC Year 2 (DLP) 2022-2023ANNIE MARGARET A/P ANTHONY MoeNo ratings yet
- Why Do Giraffes Have Long Necks?Document22 pagesWhy Do Giraffes Have Long Necks?johnosborneNo ratings yet
- Principles of Inheritance and Variations: By: Dr. Anand ManiDocument113 pagesPrinciples of Inheritance and Variations: By: Dr. Anand ManiIndu YadavNo ratings yet
- Prelim QuizDocument3 pagesPrelim QuizXyriel MacoyNo ratings yet
- Angiogenesis and Direct Myocardial RevascularizationDocument364 pagesAngiogenesis and Direct Myocardial RevascularizationPerdana SidaurukNo ratings yet
- Jagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Document305 pagesJagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Jaime Fernández-Aguirrebengoa100% (1)
- ABCD Vocal Technique Handout PDFDocument9 pagesABCD Vocal Technique Handout PDFRanieque RamosNo ratings yet
- Cell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringDocument44 pagesCell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringCrasilia Yanti PadangNo ratings yet
- Artikel SkripsiDocument12 pagesArtikel SkripsiSari RofiqohNo ratings yet
- Dihybrid Punnett SquaresDocument4 pagesDihybrid Punnett SquaresThomas Abich100% (1)
- Ovulation CalanderDocument2 pagesOvulation CalanderBN NGNo ratings yet
- Using STADEN For Sequence AssemblyDocument5 pagesUsing STADEN For Sequence AssemblyChristen Rune StensvoldNo ratings yet
- A Substitution Mutation in The Myosin Binding Protein C Gene in Ragdoll Hypertrophic CardiomyopathyDocument4 pagesA Substitution Mutation in The Myosin Binding Protein C Gene in Ragdoll Hypertrophic CardiomyopathyRoy SzeNo ratings yet
- Circulatory SystemDocument16 pagesCirculatory SystemHarinder KaurNo ratings yet
- Biosensors For Environmental ApplicationsDocument15 pagesBiosensors For Environmental ApplicationsBenjamin HonorioNo ratings yet
- The Tai Chi Code - Martial Arts Ebook by Chris DavisDocument190 pagesThe Tai Chi Code - Martial Arts Ebook by Chris DavisTomas De Wael100% (2)