You might also like
- Pericyte Biology - Novel ConceptsFrom EverandPericyte Biology - Novel ConceptsAlexander BirbrairNo ratings yet
- The Cell Cycle Inhibitor P21 Promotes The Development of - 2022 - Acta PharmaceDocument12 pagesThe Cell Cycle Inhibitor P21 Promotes The Development of - 2022 - Acta PharmaceMohammed Shuaib AhmedNo ratings yet
- s13223-022-00734-7Document8 pagess13223-022-00734-7Cher IshNo ratings yet
- 85471Document7 pages85471Vinnie Juliana YonatanNo ratings yet
- Medical InfosDocument9 pagesMedical InfosPaul AsturbiarisNo ratings yet
- Developmental and Comparative Immunology: Ying Zhang, Hui Sun, Huilin Zhao, Xingxing Chen, Jiaojiao Li, Boqing LiDocument6 pagesDevelopmental and Comparative Immunology: Ying Zhang, Hui Sun, Huilin Zhao, Xingxing Chen, Jiaojiao Li, Boqing LiMohammad Farhan AdityaNo ratings yet
- Stem Cells and Repair of Lung InjuriesDocument9 pagesStem Cells and Repair of Lung InjuriesDani PazNo ratings yet
- Eosinofilos Proteomica de Su Activacion Frontiers Medicine 2018Document8 pagesEosinofilos Proteomica de Su Activacion Frontiers Medicine 2018Gustavo GomezNo ratings yet
- New Generation of Cell-Penetrating Peptides Functionality Andpotential Clinical ApplicationDocument14 pagesNew Generation of Cell-Penetrating Peptides Functionality Andpotential Clinical ApplicationMoritz ListNo ratings yet
- Endothelium: Please Scroll Down For ArticleDocument12 pagesEndothelium: Please Scroll Down For Articleshimonl3892No ratings yet
- 09 Light Scattering Measurements of Sub Cellular Structure Providee Noninvasive Early Detection of Chemotherapy-Induced ApoptosisDocument7 pages09 Light Scattering Measurements of Sub Cellular Structure Providee Noninvasive Early Detection of Chemotherapy-Induced ApoptosisLai MattNo ratings yet
- Bone Marrow Mononuclear Cell Therapy in Experimental Allergic AsthmaDocument10 pagesBone Marrow Mononuclear Cell Therapy in Experimental Allergic Asthmayarimar hoyosNo ratings yet
- Sulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitionDocument10 pagesSulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitiongardamdNo ratings yet
- s03 - Fibrosis From Mechanisms To MedicineDocument12 pagess03 - Fibrosis From Mechanisms To MedicineWilly CaballeroNo ratings yet
- FOXK1 Plays An Oncogenic Role in The Deve 2017 Biochemical and Biophysical RDocument7 pagesFOXK1 Plays An Oncogenic Role in The Deve 2017 Biochemical and Biophysical RFlorin AchimNo ratings yet
- Enteropathogenic Escherichia Coli Infection in ChildrenDocument10 pagesEnteropathogenic Escherichia Coli Infection in ChildrenAnca MareNo ratings yet
- Distrofias Musculares 2019 MercuriDocument14 pagesDistrofias Musculares 2019 Mercurimaria palaciosNo ratings yet
- JANG - Extracellular Vesicle (EV) - Polyphenol Nanoaggregates For microRNA-based Cancer DiagnosisDocument10 pagesJANG - Extracellular Vesicle (EV) - Polyphenol Nanoaggregates For microRNA-based Cancer DiagnosisenaNo ratings yet
- Xuyen Tam Lien - Lien Phoi Sau Ton ThuongDocument9 pagesXuyen Tam Lien - Lien Phoi Sau Ton ThuongphicammienNo ratings yet
- Induced Pluripotent Stem Cell-Derived Lung Alveolar Epithelial Type II Cells Reduce Damage in Bleomycin-Induced Lung FibrosisDocument12 pagesInduced Pluripotent Stem Cell-Derived Lung Alveolar Epithelial Type II Cells Reduce Damage in Bleomycin-Induced Lung FibrosisPercy SolisNo ratings yet
- OPA1 Overexpression Ameliorates Mitochondrial Cristae Remodeling, Mitochondrial Dysfunction, and Neuronal Apoptosis in Prion DiseasesDocument13 pagesOPA1 Overexpression Ameliorates Mitochondrial Cristae Remodeling, Mitochondrial Dysfunction, and Neuronal Apoptosis in Prion DiseasesSerque777No ratings yet
- Structures of The Entire Human Opioid Receptor FamDocument33 pagesStructures of The Entire Human Opioid Receptor FamAndreea SimaNo ratings yet
- 2017년 Biomaterials xEntrDocument14 pages2017년 Biomaterials xEntrjonghyeokshin7No ratings yet
- Yoon 2014Document7 pagesYoon 2014aleegaleanoNo ratings yet
- 10.1007@s00424-019-02334-8Document16 pages10.1007@s00424-019-02334-8Martín FleiNo ratings yet
- jpm-11-00771-with-coverDocument9 pagesjpm-11-00771-with-coverutsabmondal08No ratings yet
- Artigo Sobre A Osteopontina Versão OriginalDocument5 pagesArtigo Sobre A Osteopontina Versão OriginalAjaxerrorNo ratings yet
- Alv 1Document11 pagesAlv 1Widia MeliawatiNo ratings yet
- Cao 2013Document9 pagesCao 2013Magno FilhoNo ratings yet
- ASIP Centennial Review: Cerebral MalariaDocument9 pagesASIP Centennial Review: Cerebral MalariaAndi Karwana CiptaNo ratings yet
- COPD 377026 Identification of Small Airway Epithelium Related Hub GenesDocument15 pagesCOPD 377026 Identification of Small Airway Epithelium Related Hub GenesdyahNo ratings yet
- Myeloperoxydase Expression in The Pathogenesis of Nasal PolypsDocument4 pagesMyeloperoxydase Expression in The Pathogenesis of Nasal PolypsSiq Febri SmnjtkNo ratings yet
- Apoptosis y EfermedadDocument17 pagesApoptosis y EfermedadCharles TipanNo ratings yet
- Inhibiting Collagen I Production and Tumor Cell Colonizatio - 2022 - Acta PharmaDocument13 pagesInhibiting Collagen I Production and Tumor Cell Colonizatio - 2022 - Acta PharmaMohammed Shuaib AhmedNo ratings yet
- Am 31 1902791Document32 pagesAm 31 1902791Austin TohNo ratings yet
- J Nano 2011 06 011Document10 pagesJ Nano 2011 06 011HoangNgocAnhNhanNo ratings yet
- Almeida and Wu JC 2011Document9 pagesAlmeida and Wu JC 2011Danielle SantosNo ratings yet
- Cell-Specific Retention and Action of Pheophorbide-Based Photosensitizers in Human Lung Cancer CellsDocument38 pagesCell-Specific Retention and Action of Pheophorbide-Based Photosensitizers in Human Lung Cancer Cellskamal.nayanNo ratings yet
- RETRACTED The First Tissue Engineered Airway TranDocument7 pagesRETRACTED The First Tissue Engineered Airway TranJoni SiahaanNo ratings yet
- EosinófilosDocument13 pagesEosinófilosdkbritobNo ratings yet
- Wang 2019Document10 pagesWang 2019Sebastian MilesNo ratings yet
- Erythropoietin Surprises. An Immune SagaDocument2 pagesErythropoietin Surprises. An Immune SagalmvalliNo ratings yet
- Cancer Science - 2022 - Wang - The Emerging Multifaceted Role of PINK1 in Cancer BiologyDocument11 pagesCancer Science - 2022 - Wang - The Emerging Multifaceted Role of PINK1 in Cancer BiologyT Smith AndresNo ratings yet
- From Endoplasmic-Reticulum Stress To The Inflammatory ResponseDocument22 pagesFrom Endoplasmic-Reticulum Stress To The Inflammatory ResponseivanovichNo ratings yet
- ACEL 18 E12858Document15 pagesACEL 18 E12858Valen EstevezNo ratings yet
- Review Sepsis CCM 2007Document9 pagesReview Sepsis CCM 2007Nicole MariaNo ratings yet
- Fatta Hi 2017Document3 pagesFatta Hi 2017MedranoReyesLuisinNo ratings yet
- Onconephrology 2022 An Update.19Document14 pagesOnconephrology 2022 An Update.19Prakash GudsoorkarNo ratings yet
- Proteome Profiling of Lung Tissues in ChronicDocument8 pagesProteome Profiling of Lung Tissues in ChronicAndreea MoalesNo ratings yet
- Alyu 2016Document16 pagesAlyu 2016Jessy CANo ratings yet
- Recent Advances in Physiologically Based Pharmacokinetics and Pharmacodynamic Models For Anticancer NanomedicinesDocument20 pagesRecent Advances in Physiologically Based Pharmacokinetics and Pharmacodynamic Models For Anticancer NanomedicinesEsteban MalamboNo ratings yet
- 1 s2.0 S1569199321019548 MainDocument1 page1 s2.0 S1569199321019548 MainAnita DewiNo ratings yet
- Fimmu 13 814030Document15 pagesFimmu 13 814030yunitaknNo ratings yet
- Comparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismDocument20 pagesComparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismCristina BulgaruNo ratings yet
- Quercetin alleviates acute kidney injury by inhibiting ferroptosisDocument13 pagesQuercetin alleviates acute kidney injury by inhibiting ferroptosisdiego.battiatoNo ratings yet
- Imunologi - Bella Suci Ramadhani - 42118005 - Review 2Document15 pagesImunologi - Bella Suci Ramadhani - 42118005 - Review 2BellaNo ratings yet
- Porro 2021Document9 pagesPorro 20210hitk0No ratings yet
- Journal Pre-Proof: ToxicologyDocument33 pagesJournal Pre-Proof: ToxicologyKreAch3RNo ratings yet
- A Systematic Review of Procyanidins Updates On Current Bioactivities and Potential Benefits in Wound HealingDocument6 pagesA Systematic Review of Procyanidins Updates On Current Bioactivities and Potential Benefits in Wound HealingMikail Ahmad FawzyNo ratings yet
- Advances in Stem Cells and Regenerative Medicine: Single-Cell Dynamics, New Models and Translational PerspectivesDocument5 pagesAdvances in Stem Cells and Regenerative Medicine: Single-Cell Dynamics, New Models and Translational PerspectivesVeridica El EdgieNo ratings yet
- Microrna Sequence Codes For Small Extracellular Vesicle Release and Cellular RetentionDocument28 pagesMicrorna Sequence Codes For Small Extracellular Vesicle Release and Cellular RetentionValen EstevezNo ratings yet
- Mouse Mesenchymal Stem Cell-Derived Exosomal Mir-466F-3P Reverses Emt Process Through Inhibiting Akt/Gsk3Β Pathway Via C-Met In Radiation-Induced Lung InjuryDocument16 pagesMouse Mesenchymal Stem Cell-Derived Exosomal Mir-466F-3P Reverses Emt Process Through Inhibiting Akt/Gsk3Β Pathway Via C-Met In Radiation-Induced Lung InjuryValen EstevezNo ratings yet
- An rRNA Fragment in Extracellular Vesicles Secreted by Human Airway Epithelial Cells Increases The Fluoroquinolone Sensitivity of P. AeruginosaDocument39 pagesAn rRNA Fragment in Extracellular Vesicles Secreted by Human Airway Epithelial Cells Increases The Fluoroquinolone Sensitivity of P. AeruginosaValen EstevezNo ratings yet
- TP53 Mutations Upregulate RCP Expression Via Sp1/3 To Drive Lung Cancer ProgressionDocument15 pagesTP53 Mutations Upregulate RCP Expression Via Sp1/3 To Drive Lung Cancer ProgressionValen EstevezNo ratings yet
- Microglial NF-κB drives tau spreading and toxicity in a mouse model of tauopathyDocument19 pagesMicroglial NF-κB drives tau spreading and toxicity in a mouse model of tauopathyValen EstevezNo ratings yet
- A Natural Variation-Based Screen in Mouse Cells Reveals USF2 As A Regulator of The DNA Damage Response and Cellular SenescenceDocument49 pagesA Natural Variation-Based Screen in Mouse Cells Reveals USF2 As A Regulator of The DNA Damage Response and Cellular SenescenceValen EstevezNo ratings yet
- Torn About Sasp RegulationDocument2 pagesTorn About Sasp RegulationValen EstevezNo ratings yet
- Circadian Regulation of Protein Cargo in Extracellular VesiclesDocument14 pagesCircadian Regulation of Protein Cargo in Extracellular VesiclesValen EstevezNo ratings yet
- Supplementary Figures: WT1 Ck7 WT1 Ck7Document21 pagesSupplementary Figures: WT1 Ck7 WT1 Ck7Valen EstevezNo ratings yet
- Development of iPSC-based Clinical Trial Selection Platform For Patients With Ultrarare DiseasesDocument17 pagesDevelopment of iPSC-based Clinical Trial Selection Platform For Patients With Ultrarare DiseasesValen EstevezNo ratings yet
- Cellular Signalling: Antero Salminen, Kai KaarnirantaDocument6 pagesCellular Signalling: Antero Salminen, Kai KaarnirantaValen EstevezNo ratings yet
- Sciadv Abh2445Document14 pagesSciadv Abh2445Valen EstevezNo ratings yet
- Combining Stem Cell Rejuvenation and Senescence Targeting To Synergistically Extend LifespanDocument31 pagesCombining Stem Cell Rejuvenation and Senescence Targeting To Synergistically Extend LifespanValen EstevezNo ratings yet
- Sciadv Abf8627Document18 pagesSciadv Abf8627Valen EstevezNo ratings yet
- Aging 07 0986Document14 pagesAging 07 0986Valen EstevezNo ratings yet
- Pebbles and Sand On Asteroid (162173) Ryugu: in Situ Observation and Particles Returned To EarthDocument7 pagesPebbles and Sand On Asteroid (162173) Ryugu: in Situ Observation and Particles Returned To EarthValen EstevezNo ratings yet
- Early SRC Activation Skews Cell Fate From Apoptosis To SenescenceDocument17 pagesEarly SRC Activation Skews Cell Fate From Apoptosis To SenescenceValen EstevezNo ratings yet
- Autolysosomal Degradation of Cytosolic Chromatin Fragments Antagonizes Oxidative Stress-Induced SenescenceDocument13 pagesAutolysosomal Degradation of Cytosolic Chromatin Fragments Antagonizes Oxidative Stress-Induced SenescenceValen EstevezNo ratings yet
- ACEL 18 E12858Document15 pagesACEL 18 E12858Valen EstevezNo ratings yet
- CK2 Down-Regulation Increases the Expression of Senescence-Associated Secretory Phenotype Factors through NF-κB ActivationDocument12 pagesCK2 Down-Regulation Increases the Expression of Senescence-Associated Secretory Phenotype Factors through NF-κB ActivationValen EstevezNo ratings yet
- STAT5A Modulates CDYL2/SLC7A6 Pathway To Inhibit The Proliferation and Invasion of Hepatocellular Carcinoma by Targeting To mTORC1Document13 pagesSTAT5A Modulates CDYL2/SLC7A6 Pathway To Inhibit The Proliferation and Invasion of Hepatocellular Carcinoma by Targeting To mTORC1Valen EstevezNo ratings yet
- Mirna-29A Inhibits Atherosclerotic Plaque Formation by Mediating Macrophage Autophagy Via Pi3K/Akt/Mtor PathwayDocument14 pagesMirna-29A Inhibits Atherosclerotic Plaque Formation by Mediating Macrophage Autophagy Via Pi3K/Akt/Mtor PathwayValen EstevezNo ratings yet
- Oxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationDocument13 pagesOxidative Stress-Induced Circkif18A Downregulation Impairs Mcm7-Mediated Anti-Senescence in Intervertebral Disc DegenerationValen EstevezNo ratings yet
- Cancer Letters: Original ArticlesDocument11 pagesCancer Letters: Original ArticlesValen EstevezNo ratings yet
- 3'Utr-Directed, Kinase Proximal Mrna Decay Inhibits C/Ebpβ Phosphorylation/Activation To Suppress Senescence In Tumor CellsDocument50 pages3'Utr-Directed, Kinase Proximal Mrna Decay Inhibits C/Ebpβ Phosphorylation/Activation To Suppress Senescence In Tumor CellsValen EstevezNo ratings yet
- Online Methods Cell Lines and Cell CultureDocument15 pagesOnline Methods Cell Lines and Cell CultureValen EstevezNo ratings yet
- J of Extracellular Bio - 2022 - Ramos - The Functional Role of Soluble Proteins Acquired by Extracellular VesiclesDocument21 pagesJ of Extracellular Bio - 2022 - Ramos - The Functional Role of Soluble Proteins Acquired by Extracellular VesiclesValen EstevezNo ratings yet
- Genetically Engineered Mri-Trackable Extracellular Vesicles As Sars-Cov-2 Mimetics For Mapping Ace2 Binding in VivoDocument23 pagesGenetically Engineered Mri-Trackable Extracellular Vesicles As Sars-Cov-2 Mimetics For Mapping Ace2 Binding in VivoValen EstevezNo ratings yet
- Extracellular Vesicle (EV) Mitochondria and EV/HSP27 Mixtures Protect The Ischemic Brain EndotheliumDocument71 pagesExtracellular Vesicle (EV) Mitochondria and EV/HSP27 Mixtures Protect The Ischemic Brain EndotheliumValen EstevezNo ratings yet
- Possible Mechanism of Porosome-Mediated Exosome Release: Key Terms: Neuronal Porosome, Beta CellDocument30 pagesPossible Mechanism of Porosome-Mediated Exosome Release: Key Terms: Neuronal Porosome, Beta CellValen EstevezNo ratings yet
- CardiomyopathiesDocument9 pagesCardiomyopathiesSaurabh BhandariNo ratings yet
- Worksheet Plant Growth 1 - ANSWERSDocument4 pagesWorksheet Plant Growth 1 - ANSWERSDwight DsilvaNo ratings yet
- Toxic Gas DispersionDocument13 pagesToxic Gas Dispersionsmart rusdanartoNo ratings yet
- Plant Anatomy and PhysiologyDocument29 pagesPlant Anatomy and PhysiologyAileen Mae TabileNo ratings yet
- أجزاء جسم النحلةDocument71 pagesأجزاء جسم النحلةzianiNo ratings yet
- Kimia Kofaktor Dan KoenzimDocument35 pagesKimia Kofaktor Dan KoenzimSuwahono, M.PdNo ratings yet
- Gen Bio 2Document35 pagesGen Bio 2acers filedumpNo ratings yet
- NEUROMUSCULAR TRANSMISSION: ANATOMY AND PHYSIOLOGYDocument45 pagesNEUROMUSCULAR TRANSMISSION: ANATOMY AND PHYSIOLOGYparuNo ratings yet
- Effect of Exercise On Pulse RateDocument2 pagesEffect of Exercise On Pulse RateMaya AmmarNo ratings yet
- CompleteDocument7 pagesCompleteJasmine MendozaNo ratings yet
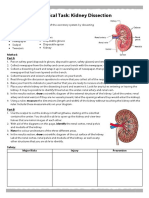
- Practical Kidney Dissection: Identify StructuresDocument3 pagesPractical Kidney Dissection: Identify StructuresIvona Matković100% (1)
- Y3 Bio Notes Unit 1Document24 pagesY3 Bio Notes Unit 1SANJAY SHIVANINo ratings yet
- Swiss Guidelines For Safe 1.7 Dry Needling 01Document21 pagesSwiss Guidelines For Safe 1.7 Dry Needling 01monkeyhsuNo ratings yet
- What Is BiologyDocument3 pagesWhat Is Biologymichellouise17No ratings yet
- Gen Bio 1 Lesson 4Document2 pagesGen Bio 1 Lesson 4Ric SantosNo ratings yet
- Laporan Bulanan Mutasi Barang Persediaan (Habis Pakai Laborat)Document16 pagesLaporan Bulanan Mutasi Barang Persediaan (Habis Pakai Laborat)putraNo ratings yet
- HUMAN ANATOMY AND PHYSIOLOGY - Cell NotesDocument9 pagesHUMAN ANATOMY AND PHYSIOLOGY - Cell NotesManish BudhrajaNo ratings yet
- What Determines Protein QualityDocument6 pagesWhat Determines Protein QualitysuprimoNo ratings yet
- Proteinuria and Hypoalbuminemia, SN CaseDocument5 pagesProteinuria and Hypoalbuminemia, SN CasekustiahNo ratings yet
- Mitochondrial oxidative phosphorylation and the proton gradientDocument47 pagesMitochondrial oxidative phosphorylation and the proton gradientSadaf BegNo ratings yet
- CHSE Science Revised Syllabus 20-21 PDFDocument106 pagesCHSE Science Revised Syllabus 20-21 PDFsandeep prASAD PATRONo ratings yet
- Fast Growing Cyanobacterial Chassis For Synthetic Biology ApplicationDocument16 pagesFast Growing Cyanobacterial Chassis For Synthetic Biology Application王礼耀100% (2)
- Scientific TimesDocument2 pagesScientific TimesNicolas PalaciosNo ratings yet
- Basic Principles of TaxonomyDocument39 pagesBasic Principles of TaxonomyRaiven Rhyle Reyes0% (1)
- KS4 Homework 02 Year 9Document2 pagesKS4 Homework 02 Year 9conquester rhanmiaNo ratings yet
- Calvin CycleDocument6 pagesCalvin CycleAtika ZulfiqarNo ratings yet
- Water Food and DieteticsDocument31 pagesWater Food and DieteticsPrecious Tinashe NyakabauNo ratings yet
- Full Blood PictureDocument1 pageFull Blood PictureGerardLumNo ratings yet
- IFU Dia TT EN 20170915Document2 pagesIFU Dia TT EN 20170915P managerNo ratings yet
- Principles of Drug ActionDocument42 pagesPrinciples of Drug ActionsharmamtechNo ratings yet