Professional Documents
Culture Documents
Effect of Obesity On High-Density Lipoprotein Metabolism
Uploaded by
Klinik FiraraOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Effect of Obesity On High-Density Lipoprotein Metabolism
Uploaded by
Klinik FiraraCopyright:
Available Formats
See discussions, stats, and author profiles for this publication at: https://www.researchgate.
net/publication/5653834
Effect of Obesity on High-density Lipoprotein Metabolism**
Article in Obesity · January 2008
DOI: 10.1038/oby.2007.342 · Source: PubMed
CITATIONS READS
117 245
2 authors, including:
Shirya Rashid
182 PUBLICATIONS 2,349 CITATIONS
SEE PROFILE
Some of the authors of this publication are also working on these related projects:
Public Health Concerns in South Asia View project
Drug Discovery and Development View project
All content following this page was uploaded by Shirya Rashid on 22 May 2019.
The user has requested enhancement of the downloaded file.
Review
Effect of Obesity on High-density Lipoprotein
Metabolism
Shirya Rashid and Jacques Genest
Abstract lipid transfer to HDL by lipoprotein lipase and reduced
RASHID, SHIRYA, JACQUES GENEST. Effect of obesity HDL clearance by hepatic triglyceride lipase as a result of
on high-density lipoprotein metabolism. Obesity. 2007;15: endurance training are two important mechanisms for in-
2875–2888. creases in HDL observed from exercise.
Reduced levels of high-density lipoproteins (HDL) in non-
obese and obese states are associated with increased risk for Key words: adiposity, high-density lipoproteins, diet,
the development of coronary artery disease. Therefore, it is exercise, cardiovascular risk
imperative to determine the mechanisms responsible for
reduced HDL in obese states and, conversely, to examine
therapies aimed at increasing HDL levels in these individ-
Introduction
The strong inverse relationship between plasma concen-
uals. This paper examines the multiple causes for reduced
trations of high-density lipoprotein (HDL)1-cholesterol and
HDL in obese states and the effect of exercise and diet—
the risk of developing coronary artery disease (CAD) (1)
two non-pharmacologic therapies— on HDL metabolism in
has stimulated interest in determining the mechanisms and
humans. In general, the concentration of HDL-cholesterol is
optimal management of low HDL-cholesterol (⬍35 mg/dL)
adversely altered in obesity, with HDL-cholesterol levels
(2,3). A number of genetic, lifestyle, and environmental
associated with both the degree and distribution of obesity.
factors have been shown to contribute to the lowering of
More specifically, intra-abdominal visceral fat deposition is HDL-cholesterol. Genetic disorders characterized by low
an important negative correlate of HDL-cholesterol. The HDL-cholesterol concentrations are rare and include
specific subfractions of HDL that are altered in obese states mutations in apolipoprotein A-I (4 – 6), the major protein
include the HDL2, apolipoprotein A-I, and pre-1 subfrac- component of HDL, defects in HDL-mediated efflux of
tions. Decreased HDL levels in obesity have been attributed cholesterol from peripheral cells through the membrane
to both an enhancement in the uptake of HDL2 by adipo- transporter ABCA1 (7–9), and hypercatabolism of normal
cytes and an increase in the catabolism of apolipoprotein apolipoprotein A-I (10). In contrast, lifestyle factors—
A-I on HDL particles. In addition, there is a decrease in the including obesity, physical inactivity, diet, smoking, and
conversion of the pre-1 subfraction, the initial acceptor of advanced alcohol-related liver disease—account for the
cholesterol from peripheral cells, to pre-2 particles. Con- majority of cases of low HDL-cholesterol levels (11–13).
versely, as a means of reversing the decrease in HDL levels In particular, obesity, defined as the excessive accumu-
in obesity, sustained weight loss is an effective method. lation of body fat, is frequently associated with a low
More specifically, weight loss achieved through exercise is concentration, adverse distribution pattern, and abnormal
more effective at raising HDL levels than dieting. Exercise metabolism of HDL particles (14,15). Obesity is a highly
mediates positive effects on HDL levels at least partly prevalent chronic state observed in 32% of all adults in the
through changes in enzymes of HDL metabolism. Increased United States (16). Moreover, the Framingham Heart Study
(17) and the Third National Health and Nutrition Examina-
tion Survey (18) have shown a significant increase in life-
Received for review January 17, 2007.
Accepted in final form May 3, 2007.
time risk of CAD with level of obesity, which may be partly
The costs of publication of this article were defrayed, in part, by the payment of page attributed to its association with low HDL-cholesterol lev-
charges. This article must, therefore, be hereby marked “advertisement” in accordance with
18 U.S.C. Section 1734 solely to indicate this fact.
Faculty of Medicine, Division of Cardiology, McGill University, Montreal, Canada.
1
Address correspondence to Shirya Rashid, Department of Medicine, Division of Cardiology, Nonstandard abbreviations: HDL, high-density lipoprotein; CAD, coronary artery disease;
McGill University, Royal Victoria Hospital, Room H7–13, 687 Pine Avenue West, Mon- LDL, low-density lipoprotein; VAT, visceral adipose tissue; VLDL, very-low-density li-
treal, Quebec, Canada H3A 1A1. poprotein; LCAT, lecithin cholesterol ester transfer protein; CETP, cholesteryl ester transfer
E-mail: shirya2004@yahoo.ca protein; FFA, free fatty acid; HSL, hormone-sensitive lipase; PLPT, phospholipid transfer
Copyright © 2007 NAASO protein; apo A-I, apolipoprotein A-I.
OBESITY Vol. 15 No. 12 December 2007 2875
Obesity and High-density Lipoproteins, Rashid and Genest
els. Conversely, an increase in HDL-cholesterol levels has levels both significantly declined in a linear fashion with
been proven to be cardioprotective, such that each 1-mg/dL increasing BMI (ranging from ⬍21 to ⱖ30 kg/m2) (21). An
elevation in HDL reduces CAD risk by 2% to 3%, indepen- increase in BMI, moreover, was strongly and significantly
dently of the levels of low-density lipoprotein and triglyc- related to reduced HDL levels and was more strongly re-
erides (19). A comprehensive pathophysiological explana- lated to reduced HDL than other cardiovascular risk factors,
tion for the association between obesity and increased CAD such as low-density lipoprotein (LDL)-cholesterol levels (21).
risk is still lacking. However, because the variation in Although the Quetelet index and BMI have been used as
HDL-cholesterol levels is an important modifiable CAD a measure of adiposity in many epidemiologic studies
risk factor in obese states, it is imperative to elucidate the (17,22), they are indirect indices of obesity, measuring both
mechanisms which contribute to the lowering of HDL- lean and adipose tissue mass, and they do not provide
cholesterol in obesity and to determine the efficacy of information on the distribution of body fat (23). This is
treatments aimed at raising HDL-cholesterol levels in obese particularly of concern in women, because compared with
individuals. men, the prevalence of complications in lipid metabolism
This paper will outline the evidence linking obesity to and other cardiovascular disease risk factors is less consis-
variations in HDL-cholesterol metabolism, characterize the tently correlated with the degree of obesity in women.
pathophysiologic mechanisms responsible, and examine the Instead, the distribution of body fat in women (and men) is
efficacy of weight loss achieved with exercise—the recom-
an independent predictor of the adverse metabolic patterns
mended non-pharmacologic intervention for increasing
of obesity, cardiovascular morbidity, and mortality (23,24).
HDL-cholesterol levels in obese individuals— on poten-
More specifically, studies that have examined regional fat
tially reversing these changes.
distribution, using both indirect anthropometric parameters
(waist-to-hip circumference, subcutaneous skinfold thickness)
(25) and direct measurement of adipose tissue surface areas
Evidence that Adiposity Is Associated with (computed tomography) (23), have shown a close correlation
Reduced HDL-cholesterol Levels between intra-abdominal fat deposition and indices of car-
Several indices of adiposity have been used to show an diovascular risk, including a low HDL-cholesterol/total cho-
association between obesity and HDL-cholesterol. Epide- lesterol ratio, hypertension, glucose intolerance, hypertri-
miologic data from the large, cross-sectional multinational glyceridemia, and hyperinsulinemia (23,25,26). Such an
Lipid Research Clinics Program Prevalence Study found a accumulation of intra-abdominal visceral fat is more charac-
significant inverse correlation between the Quetelet index of teristic of men, who generally exhibit android or upper body
body mass (height divided by weight squared) and total obesity, than of women, who more typically exhibit gynoid or
HDL-cholesterol in adults of both sexes (20). The associa- lower body obesity, and explains the greater correlation of total
tion between the Quetelet index and HDL-cholesterol re-
fat distribution to an abnormal metabolic profile in men (27).
mained significant when multivariate analysis was applied
Peiris et al. (23) quantified the effect of regional fat
to control for some confounding variables affecting HDL-
deposition on HDL levels in healthy premenopausal women
cholesterol (smoking, alcohol intake, exogenous steroid
with a wide range of body weight and adiposity and found
hormone use, and plasma triglyceride levels), although oth-
(using computed tomography) that 15% of the variance in
ers were not considered (diet, habitual exercise patterns). To
the HDL-cholesterol/total cholesterol ratio was explained
quantify the association, covariance-adjusted mean HDL-
cholesterol levels were calculated at the 10th and 90th by variations in visceral fat mass. However, despite the
percentiles for Quetelet index and revealed that mean HDL- significant effect of adipose tissue distribution on variations
cholesterol levels were 6 to 7 mg/dL higher at the Quetelet in HDL-cholesterol levels, body fat distribution per se has
90th percentile than at the 10th percentile. This difference in been found to be insufficient to explain observed associa-
HDL-cholesterol levels has been reported to significantly tions between visceral intra-abdominal adipose tissue and
alter the risk of mortality from CAD (1). HDL-cholesterol alterations in premenopausal women. De-
BMI, expressed as body weight in kilograms divided by spres et al. (15) showed that obesity is a necessary condition
height in square meters, is frequently used as a measure of for the association in premenopausal women. In a sample of
body fatness in large epidemiologic studies (21). The World 22 lean women with a mean BMI of 21 kg/m2, they found
Health Organization (2002) delineates individuals with a no significant correlation between waist-to-hip ratio and
BMI ⱖ25 kg/m2 and ⱕ29.9 kg/m2 as overweight and those plasma HDL levels, whereas computer tomography-mea-
with a BMI ⱖ30 kg/m2 as obese. In one large-scale study sured high trunk abdominal fat and abdominal cell hyper-
(1566 men and 1627 women) of participants from the Fra- trophy correlated negatively with plasma HDL apoliprotein
mingham Offspring Study, lipoprotein and BMI measures A-I (the major protein component of HDL particles) and
were taken to determine the effects of BMI on coronary risk HDL2 subfraction cholesterol levels (r ⫽ ⫺0.51 and p ⬍
factors. Plasma HDL-cholesterol and apolipoprotein A-1 0.05 for both).
2876 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
In postmenopausal women, in contrast, it seems that pendent predictor of HDL-cholesterol levels after adjusting
obesity is not required to be present for correlations between for BMI and waist circumference, independently of age and
intra-abdominal fat and HDL levels. Ostlund et al. (25) height (30).
determined that variations in abdominal adiposity, as as- To our knowledge, no epidemiologic or clinical study to
sessed through the waist-to-hip ratio, explained 32% of the date has established a causal link between the low plasma
variance of HDL2-cholesterol in healthy, non-obese (or HDL-cholesterol concentrations observed in obese states
mildly obese) elderly (60 to 70 years of age) men and and an increased risk for development of CAD events. Other
women (independently of sex). It has been found both in lipoprotein abnormalities, such as increases in the athero-
perimenopausal and postmenopausal obese women that a genic very-low-density lipoproteins (VLDLs) and LDLs,
high visceral adipose tissue (VAT) area, as assessed with and other metabolic alterations, such as glucose intolerance
computer tomography, is associated with low HDL concen- and insulin resistance, are also associated with obesity and
trations (28). Women with VAT of 106 to 162 cm2 are 2.5 likely contribute to the increased CAD risk in obesity. These
times more likely to have low HDL-cholesterol levels, and changes, however, seem to occur less consistently than low
women with a VAT of ⱖ163 cm2 are 5.5 times more likely HDL-cholesterol levels in obesity. Moreover, the knowl-
to have low HDL-cholesterol, than women with a VAT of edge that HDL-cholesterol levels are an important marker of
105 cm2 or less (28). Thus, the appropriate index of adi- CAD risk and the finding that low HDL-cholesterol levels
are consistently associated with several indices of adiposity
posity to apply in studies investigating associations between
have stimulated research on the mechanisms of HDL low-
adiposity and plasma HDL-cholesterol concentrations is
ering in obese states. These mechanisms will be reviewed in
dependent on the age, metabolic state, and sex of the par-
the next section.
ticular subpopulation/sample group under study.
Studies have additionally examined the impact of ethnic-
ity on HDL levels and whether adiposity contributes to the
relationship. For example, compared with white individuals,
Mechanisms of HDL Lowering in Obesity:
black individuals have consistently been characterized by
Effect of Obesity on HDL Subfractions
In addition to the levels of HDL, the distribution and
higher plasma HDL-cholesterol concentrations. In one
composition of HDL subfractions determine their functional
study, metabolic risk variables, including plasma HDL-
effectiveness. HDL particles are a discrete but highly het-
cholesterol levels, along with body fatness and VAT accu- erogeneous group of lipoproteins that differ in density, size,
mulation, were measured in healthy white men and women lipid content, and protein composition (31). Not surpris-
and black (African-American) men and women in North ingly, obesity has been shown to adversely affect the levels
America (29). Plasma HDL-cholesterol levels were found to of HDL subfractions principally associated with cardiopro-
be higher in black men and women than in their white tection: HDL2, apolipoprotein A-I, and pre-1. This section
counterparts (29). This elevation in HDL-cholesterol levels will examine the mechanisms by which obesity is reported
could not be explained by the extent of subcutaneous fat or to mediate variations in HDL subfractions and their impli-
total body fat mass in the black individuals vs. the white cations.
individuals, because no race differences were found in these
variables (29). However, a lower accumulation of VAT in
Effect of Obesity on HDL2 Levels
blacks vs. whites accompanied their higher plasma HDL-
There are several classification systems that have been
cholesterol levels and, indeed, could explain, at least in part, used to characterize HDL subpopulations. Early studies on
their elevated HDL-cholesterol levels (29). In contrast, eth- the effect of obesity on HDL subfractions used the tradi-
nicity per se provided only a minor contribution to the tional sequential ultracentrifugation and precipitation tech-
variance in plasma HDL-cholesterol levels (29). niques to separate HDL into its two principle subfractions:
Although in the case of black individuals, compared with the less dense, cholesterol- and phospholipid-enriched
white individuals, differences in adiposity contribute to HDL2 particles and the denser, smaller, and relatively pro-
differences in HDL-cholesterol levels, this is not always the tein rich HDL3 fraction (2). HDL2 particles are generated
case when comparing different ethnic groups. In another from HDL3 particles when triglyceride-rich lipoproteins are
study, healthy men and women of European and South hydrolyzed by lipoprotein lipase and subsequently transfer
Asian descent in North America were assessed for their their surface components (including cholesterol and phos-
anthropometric variables and lipid levels (30). Although pholipid) to HDL3 (2). HDL2 particles are also formed from
BMI and waist circumference were similar between Euro- smaller HDL particles when lecithin cholesterol ester trans-
pean and South Asian men and European and South Asian fer protein (LCAT) esterifies cholesterol within HDL (32).
women, South Asian men and women had significantly Conversely, the smaller, denser HDL3 subfraction is formed
lower HDL-cholesterol values compared with their Euro- when HDL2 particles are hydrolyzed by the enzyme hepatic
pean counterparts (30). In this case, ethnicity was an inde- lipase.
OBESITY Vol. 15 No. 12 December 2007 2877
Obesity and High-density Lipoproteins, Rashid and Genest
Because of the significant cardioprotective role of HDL2,
the concentrations of HDL2-cholesterol and its relationship
to obesity were studied in a number of different sample
population groups. In one study comparing HDL concen-
trations in 68 age-matched obese (mean BMI ⫽ 32 ⫾ 4
kg/m2) and control subjects, cholesterol and protein con-
tents of HDL2 were found to be 50% lower in the obese
subjects (33). In addition, there was a greater negative
correlation between BMI and HDL2 than between BMI and
HDL3 levels. It was believed that the reduced HDL2 levels
in the obese subjects were caused by two factors: an overall
elevation in very-low-density triglycerides and a slight in-
crease in glycemia, reflecting insulin resistance (33). These
factors may have contributed to decreased lipolysis and a
defective transfer of surface elements to HDL3 in the obese
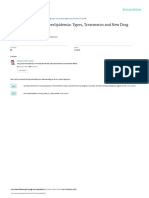
subjects (33). Figure 1: CETP mediates the transfer of triglycerides from the
Other studies conducted in obese premenopausal women triglyceride-rich lipoproteins— chylomicrons and VLDLs—to
(26) and older adults of both sexes of varying adiposity (25) HDLs. CETP also mediates the reverse transfer of cholesteryl ester
were designed to investigate the importance of adipose from the core of HDL particles to VLDLs and chylomicrons. The
tissue distribution on HDL2-cholesterol levels. These stud- net result is the formation of large, triglyceride-rich, cholesterol
ies showed in their respective sample populations that intra- ester core– depleted HDL particles, which are an ideal substrate for
abdominal fat was a more important independent predictor hepatic lipase, that are cleared by the liver.
of low HDL2-cholesterol levels than mid-thigh fat (26),
BMI, or total percentage body fat (25).
In another study, examining the contribution of intra- subjects (37). In hypertriglyceridemic states, such as obe-
abdominal fat to sex differences in hepatic lipase activity sity, the plasma protein, cholesteryl ester transfer protein
and HDL heterogeneity, it was determined that intra-ab- (CETP), mediates a greater net transfer of triglycerides
dominal fat is a major determinant of the sex differences in from the triglyceride-rich lipoproteins (VLDLs and chylo-
hepatic lipase activity (34). Men have significantly more microns) to HDL particles than normal (38,39) (Figure 1).
intra-abdominal fat and a higher hepatic lipase activity and This has been shown to produce triglyceride-rich HDL
those result in lower HDL2 levels in men (34). particles, including HDL2 particles (38). Triglyceride-rich
Data from several cross-sectional studies conducted HDL particles are also formed from a greater pool of VLDL
among survivors of myocardial infarction and among pa- particles in hypertriglyceridemic, insulin-resistant states. In
tients with angiographically-measured atherosclerosis (35) some cases of hypertriglyceridemia, insulin resistance is
indicated that levels of both HDL2 and HDL3 subfractions present, and this insulin resistance then drives the release of
were significantly lower in these subjects than in healthy free fatty acids (FFAs) from adipose tissue to the circulation
control subjects. However, the differences were greater for (40). This feed-forward cycle of FFA release into the cir-
HDL2 and especially for the relatively rich, large HDL2 culation is mediated by the enzyme hormone-sensitive
species, HDL2b. Other studies have determined that HDL2 is lipase (HSL) (41,42). HAL activity is normally suppressed
the most reliable and sensitive predictor of a normolipi- by insulin after a meal (41,42). However, the activity of this
demic individual’s ability to clear atherogenic triglyceride- lipase is enhanced in insulin-resistant states, such as obesity,
rich lipoproteins from plasma postprandially (36). These driving the release of FFAs into the circulation (41,42). The
studies suggest, but do not prove, that HDL2 vs. HDL3 FFAs provide the substrate for the formation of VLDL
levels are more closely associated with protection against particles by the liver (40). The result is a greater mass of
CAD. Conversely, they suggest that the reduction in HDL2 VLDL particles in the circulation from which a greater
levels in obese states may be associated with increased risk amount of triglycerides are transferred to HDL particles,
of CAD events. causing the HDL particles to be triglyceride rich (40).
The possible mechanisms by which obesity promotes Triglyceride-rich, large HDL2 particles are the preferred
reduced HDL2-cholesterol levels may be divided into indi- substrate for the enzyme hepatic lipase, which hydrolyzes
rect and direct processes. First, other metabolic abnor- HDL and promotes hepatic HDL uptake (38,43). Because
malities present in obese states may indirectly cause the obese subjects have been found to have significantly in-
lowering of HDL2 levels in obese individuals. Hyper- creased protein mass and activities of CETP and post-
triglyceridemia, in particular, is frequently associated with heparin hepatic lipase (44), triglyceride enrichment of
reduced HDL levels and enhanced HDL catabolism in obese HDL2 by CETP and VLDL and its subsequent hydrolysis by
2878 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
hepatic lipase may be a potential mechanism by which an elevation in endothelial lipase with increasing visceral
HDL2 levels are indirectly reduced in obese hypertriglycer- adiposity but no correlation between endothelial lipase and
idemic individuals. either total HDL- or HDL2-cholesterol levels (57). Thus,
Plasma triglyceride levels, however, vary greatly in obese endothelial lipase was found not to mediate the decrease in
individuals, and a large proportion of obese individuals are HDL2 levels seen in obesity (57).
not hypertriglyceridemic (45). Moreover, significant in- The expression levels of another recently discovered
creases in HDL concentration have been shown in severely lipase has been studied in the context of obesity. The lipase
obese men and women after weight loss through dieting and —adipose triglyceride lipase—mediates the hydrolysis of
exercise, despite the lack of change in plasma triglyceride triglyceride stores in adipocytes to form FFAs, which are
levels (46). Therefore, alternate mechanisms likely play a released into the circulation (58,59). Adipose triglyceride
role in mediating reduced HDL2 levels in obese subjects. lipase, together with HSL, is important for the hydrolysis of
Because increased CETP mass and activity levels, which triglycerides in adipose tissue in basal conditions (60).
can potentially decrease HDL2 levels (as mentioned above), In a recent study, adipose tissue biopsies were taken from
have been identified in obese states (44), pharmacologic obese subjects with a wide range of insulin resistance to
inhibition of CETP with CETP inhibitors, such as the re- assess mRNA and protein levels of adipocyte triglyceride
cently characterized torcetrapib series of CETP inhibitors lipase (61). There were strong associations, independent of
(47,48), could potentially benefit obese individuals with low fat mass, between insulin resistance and hyperinsulin-
HDL2 levels. In healthy individuals, CETP mass does not emia and the mRNA and protein of adipocyte triglyceride
correlate with HDL2 (49) or total HDL (50) levels, unless lipase (61). Overall, mRNA and protein levels of adipocyte
the individuals are also hypertriglyceridemic (50) or insulin triglyceride lipase were found to be reduced in insulin-
resistant (51). Thus, CETP inhibition could potentially ben- resistant compared with insulin-sensitive subjects (61).
efit HDL metabolism in obese individuals more than in These results suggest that in insulin-resistant, obese
normal, healthy individuals.
states, decreased expression of adipose triglyceride lipase
It should be noted that a large Phase III clinical trial
decreases the mobility of FFAs from their triglyceride stores
testing the safety and efficacy of torcetrapib was recently
in adipocytes. This indicates less FFA flux from adipocytes
halted because of the higher mortality rates detected in
to the liver. Therefore, less FFA substrate would be avail-
post-cardiac event patients using torcetrapib (52,53). Thus,
able for VLDL production in the liver. With less VLDL
torcetrapib, designed as a novel drug therapy aimed at
produced, there would be less transfer of triglycerides from
raising HDL-cholesterol and reducing atherosclerosis and
VLDL to HDL (including HDL2 particles) that would make
cardiac events, has been deemed a failure. Its failure shows
the HDL particles triglyceride-rich and unstable after action
that factors other than total HDL-cholesterol levels are
by hepatic lipase. Thus, decreased expression of adipose
important to assess when evaluating pharmacologic ap-
proaches to raising HDL-cholesterol in obese patients (53). triglyceride lipase in obese, insulin-resistant states cannot
These factors include HDL particle size and composition account for reduced HDL (and HDL2) levels in these states.
(53). Moreover, the appropriate clinical endpoints in deter- The role of the protein phospholipid transfer protein
mining the efficacy of pharmaceutical approaches to raising (PLTP) has also been studied in terms of its effect on
HDL in obese (and non-obese) patients are decreasing ath- remodeling HDL particles and its activity in obese states.
erosclerosis and decreasing the vulnerability of atheroscle- PLTP has been found in vitro to remodel HDL3 particles
rotic plaques to rupture (53). (particle diameter, 8.7 nm) into two distinct populations of
Despite the failure of torcetrapib, decreasing CETP- HDL— one larger, of particle diameter 10.9 nm, and one
mediated transfer of cholesteryl ester from HDL still seems smaller, of particle diameter 7.8 nm (62). PLTP has also
to be a sound strategy for increasing HDL levels and low- been shown to alter HDL2 particles in vitro such that a
ering CAD risk (53,54) in obese (and non-obese) subjects. population of pre--like HDL particles are produced (63).
However, drugs that modify HDL structure in a different PLTP activity was found to be elevated in a group of 8
manner than torcetrapib are likely required (53). The design healthy obese men (BMI ⬎ 27 kg/m2), relative to 24 non-
of such drugs will be greatly aided by the newly character- obese individuals (64) (Figure 2). PLTP activity, moreover,
ized crystal structure of CETP (53,55). was positively related to BMI and waist-to-hip circumfer-
The role in humans of endothelial lipase, a lipase shown ence ratio (64). Conversely, weight loss with gastric bypass
to mediate a decrease in HDL levels in animal models, has surgery in severely obese middle-age women resulted in a
also been studied. Endothelial lipase has been shown to significant decrease in PLTP activity and, concomitantly, an
hydrolyze phospholipids on the surface of HDL particles increase in the size of HDL2 particles (65). Thus, a reduc-
containing either apo A-I only or both apo A-I and apo A-II tion in PLTP activity with weight loss reversed the athero-
together (56). The result is a decrease in particle size with genic small, dense HDL particles seen in obese individuals
no dissociation of the apolipoproteins (56). Studies showed (65).
OBESITY Vol. 15 No. 12 December 2007 2879
Obesity and High-density Lipoproteins, Rashid and Genest
Effect of Obesity on Apolipoprotein A-I Levels
The HDL2 subfraction contains a relatively high concen-
tration of apolipoprotein A-I (apo A-I) (70), the major
protein component of HDL contributing to ⬃50% of its
mass (39) and an important determinant of its structure and
number in the circulation. Apo A-I is a key marker of
HDL-cholesterol levels, and studies have shown that the
level of apo A-I is more closely correlated with the reduced
risk of atherosclerosis than other markers of HDL-choles-
terol (71,72).
As with levels of HDL2, apo A-I levels are frequently low
in obese patients. Obesity was shown to be associated with
reduced plasma HDL apo A-I levels in the Framingham
Offspring Study conducted in 4260 young adult men and
women (73). Both in vitro (74,75) and in vivo studies in
transgenic animals (76) have suggested that apo A-I directly
protects against the development of atherogenesis by either
preventing the oxidation or self-aggregation of proathero-
genic LDL particles within the arterial wall (75) or by
stimulating the mobilization of cholesterol from the arterial
wall (74). Therefore, the reduced apo A-I levels associated
Figure 2: PLTP mediates the transfer of phospholipids to HDL with obesity may contribute to the increased risk of CAD in
particles. PLTP action results in the formation of larger and obese subjects.
smaller HDL particles, along with the formation of pre--HDL
Brinton et al. (77), Gylling et al. (78), and Ooi et al. (79)
particles, which are cleared by the kidneys.
studied the factors responsible for low plasma HDL apo A-I
concentrations in obese subjects, using a kinetic approach.
A number of studies have suggested that obesity per se More specifically, in their studies, radioisotopes were used
directly induces a reduction in HDL-cholesterol levels. The to trace the kinetic behavior of apo A-I in a group of
factors responsible for the well-characterized exercise- normolipidemic, non-smoking obese individuals (mean
mediated rise in HDL-cholesterol levels were examined by BMI ⫽ 30 kg/m2). Both the rate of input of apo A-I into
Williams et al. (66). Post hoc and regression analysis of 64 plasma and the time that apo A-I resides in plasma have
middle-age men who had completed either a controlled been shown to influence apo A-I concentrations under dif-
moderate exercise training program for 1 year or maintained ferent metabolic conditions (80,81). In obese subjects, two
a sedentary lifestyle revealed that the majority of plasma of the above investigators (77,78) found that a reduction in
HDL2-cholesterol increase induced by moderate exercise the residence time of apo A-I in plasma—a 30% reduction
was caused by a change in body weight. Similarly, Wolf and vs. control subjects—was the principle cause of low apo A-I
Grundy (67) found that caloric restriction did not induce a levels. Conversely, the input rates of apo A-I were not
rise in HDL-cholesterol levels in obese patients until after different between obese subjects and non-obese normolipi-
most of their excess adipose tissue had been removed. demic, control subjects. Ooi et al. found that both the
These clinical studies are consistent with in vitro evi- fractional catabolic rate and production rate of apo A-I are
dence that adipose tissue can specifically bind to and me- elevated in obese individuals compared with lean controls
diate the uptake of 125I-HDL2 and HDL3 (68,69). The (79).
uptake of HDL2 by adipose tissue may be characterized as Together, these studies suggest that the obese state di-
being tissue specific, because abdominal subcutaneous adi- rectly enhances the rate of clearance and can enhance the
pocytes mediate a greater uptake and binding than omental secretion rate of HDL (measured as apo A-I) into plasma.
adipocytes (68,69). Also, HDL2 uptake by adipocytes is This was confirmed by a later study in which the level of
dependent on fat cell size, with hypertrophied cells display- intra-abdominal fat, as measured by magnetic resonance
ing greater uptake (69). imaging, strongly correlated with the clearance rate of HDL
Overall, both direct and indirect mechanisms related to apo A-I (r ⫽ 0.98, p ⫽ 0.003) (82). It is possible that the
the metabolic consequences of obesity may account for increased plasma clearance rate of apo A-I in obese subjects
reduced HDL2 levels in obese subjects. The relative impor- contributes to the increased risk of CAD in obesity.
tance of each seems to be dependent on the primary meta- The enhancement in HDL catabolism in obese states may
bolic abnormality present and the extent and distribution of be attributed to enhanced hepatic lipase and CETP activity
adiposity in the individual. levels in obesity. First, it is well established that hepatic
2880 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
lipase activity levels increase in obese states (83,84). High and an organized, i.e., thermostable, particle with two apo
hepatic lipase activity is associated with reduced levels of A-I molecules and a phospholipid disk (88). The major
HDL2 (84). Hepatic lipase may mediate this effect on HDL2 acceptors of cellular cholesterol are lipid-free apo A-I and
levels by hydrolyzing the surface triglycerides on HDL nascent ␣-LpAI (88).
particles, rendering the particles less stable, thus enhancing Sasahara et al. (87) found that the distribution of ␣-HDL
the clearance of HDL from the circulation (85). Further- and -HDL particles was shifted to smaller species in obese
more, CETP may also play a role in the enhanced clearance subjects (23 middle-age men and postmenopausal women
of HDL in obese states. CETP activity has been found to with a BMI between 25 and 36 kg/m2) compared with lean
increase significantly in obese subjects relative to control subjects (BMI ⬍ 25 kg/m2). Total HDL-cholesterol, HDL
subjects (83). Moreover, CETP activity, independent of apo A-I, and ␣1-HDL levels were significantly reduced in
post-heparin hepatic lipase and lipoprotein lipase activities, obese subjects, confirming earlier studies. However, the
has been found to correlate negatively with HDL-choles- demonstration of increased pre-1-HDL in obese subjects
terol levels (44). CETP acts to transfer triglycerides from was a novel finding. Furthermore, on both univariate and
triglyceride-rich VLDL particles and chylomicrons to HDL stepwise regression analysis, BMI was directly correlated
particles and also mediates the reverse transfer of cho- with pre-1-HDL and inversely with ␣1-HDL.
lesteryl ester from HDL to VLDL and chylomicrons (86). Possible causes of raised pre-1-HDL levels in obesity
This results in triglyceride-rich HDL particles that are de- are greater hepatic lipase and CETP activities found in
pleted of core cholesteryl ester, directly resulting in reduced obese individuals in comparison with lean controls (44).
HDL-cholesterol levels (86). Triglyceride-rich HDL parti- The origins of pre--HDL provide insight into the impli-
cles produced through CETP action are also a good sub- cations of such raised hepatic lipase and CETP activities
strate for hepatic lipase, the action of which enhances the in obesity. Pre-1-HDL are discoidal nascent HDL par-
clearance of HDL particles (86). ticles that are generated partly from excess surface lipid
and apoA-I components of HDL2 when HDL2 is catabo-
Effect of Obesity on Pre-1 Levels lized. This process occurs when HDL2 transfers cholesteryl
Reduced HDL2 cholesterol and apo A-I levels have been esters to triglyceride-rich lipoproteins through the action of
widely identified as independent CAD risk factors in obe- CETP or is hydrolyzed through the action of hepatic lipase
sity. More recently, however, a deficiency in another HDL (Figure 2). Thus, increased CETP and hepatic lipase activ-
subfraction—pre--HDL— has been identified in obese ities in obese individuals would tend to reduce ␣1- and
states. Pre--HDL is more closely linked to the primary increase 1-HDL levels.
anti-atherosclerotic function of HDL, which is mediating In addition, PLTP likely has a role in the generation of
reverse cholesterol transport. Reverse cholesterol transport pre-1 particles in obese individuals. PLTP mediates an
is the process by which cholesterol in peripheral (non- increase in phospholipids onto the surface of HDL particles,
hepatic) tissues is transported to the liver for re-use or bile resulting in a displacement of apo A-I and the generation of
acid synthesis (39). In this reverse cholesterol transport new pre-1 particles (92) (Figure 2). In humans with a wide
system, the pre--HDL subfraction is believed to promote range of BMI (19 to 43 kg/m2), PLTP activity was shown to
the efflux of cholesterol from peripheral cells (the first step be associated with BMI, body fat mass, and subcutaneous
in reverse cholesterol transport) (87) and may, conse- fat area (92). Overall, PLTP activity was significantly
quently, prevent the accumulation of cholesterol, foam higher in obese vs. non-obese individuals (92). Moreover, in
cells, and fatty lesions in cells of the arterial intima. linear regression analysis, BMI was the sole predictor of
Pre--HDL species are effectively isolated by two- plasma PLTP activity (92).
dimensional non-denaturing electrophoresis of apoA-I- Interestingly, there was also a reduction in the pre-1-
containing HDL (87– 89). Kinetic and pulse-chase experi- HDL-to-pre-2-HDL ratio in obese subjects in the above
ments conducted in fibroblasts radiolabeled with cholesterol study, suggesting an inhibition in the conversion of pre-1
and plasma suggest that three different pre--HDL subspe- to pre-2 (87). This may reflect a disruption in reverse
cies are involved in the transport of cell-derived cholesterol cholesterol transport along the HDL particle subfractions
in plasma (90,91). In this model, pre-1-HDLs are the initial (87). Indeed, in a later in vitro cell culture study by Sasahara
mediators of cholesterol efflux from peripheral cells. Sub- et al. (93), obese individuals showed an overall decrease in
sequently, they uni-directionally transfer their cholesterol the capacity of HDL subfractions to promote cholesterol
through CETP to larger pre-2-HDLs, and then pre-3- efflux from fibroblasts. Compared with lean subjects, the
HDLs, and finally to ␣-HDL (identical to HDL2), where the cholesterol efflux activity of pre-1-HDL was higher and
cholesterol is esterified by LCAT. that of pre-2-HDL was lower in obese individuals. This is
Pre- LpAI is a partially lipidated single apo A-I moiety consistent with a disturbance in the sequential transfer of
with unknown function (88). It is likely a thermodynami- cholesterol from peripheral cells to HDL subfractions in
cally unstable transition form between lipid-free apo A-I reverse cholesterol transport. Furthermore, because pre-1-
OBESITY Vol. 15 No. 12 December 2007 2881
Obesity and High-density Lipoproteins, Rashid and Genest
HDL concentrations are increased in both obesity and CAD circumference, has been found in major clinical trials (95) to
(94), it may be a link between obesity, lowered reverse consistently increase HDL-cholesterol concentrations, when
cholesterol transport, and increased risk of CAD in obese given at a dose of 20 mg/d. Rimonabant’s action in reducing
states. food intake seems to be both centrally and peripherally
mediated (95). The peripheral action is potentially through
enhanced thermogenesis through increased oxygen con-
Effect of Weight Loss on Plasma sumption in skeletal muscle and reduced hepatic and adi-
pocyte lipogenesis, among a variety of other mechanisms.
HDL-cholesterol
In the above sections, we established 1) that the levels of In the Rimonabant In Obesity–North America multi-center,
HDL-cholesterol and composition of HDL particles are randomized, double blind trial of more than 3000 obese or
adversely altered in obese individuals; 2) that the extent of overweight individuals (BMI ⱖ 30 kg/m2 and BMI ⱖ 27
kg/m2, respectively), patients were randomized to be on a
the variation is determined by the regional distribution of fat
diet (600 kcal/d deficit) and receive placebo, 5 mg/d rimon-
and specific population under study; and 3) that the alter-
abant, or 20 mg/d rimonabant (100). After 1 year, the group
ations in HDL can lead to a reduction in the cardioprotective
receiving 20 mg/d rimonabant achieved greater mean reduc-
functions of HDL. This section will focus mainly on the
tions in weight and waist circumference and a greater in-
mechanisms and efficacy of weight loss achieved non-
crease in HDL-cholesterol than the placebo group (100). It
pharmacologically through exercise in raising HDL-choles-
has been estimated that ⬃47% to 58% of the improvement
terol levels in overweight individuals.
in HDL-cholesterol mediated by rimonabant is beyond that
As a brief explanation of the pharmacologic weight loss
expected through weight loss alone and may be mediated by
options, orlistat, sibutramine, and rimonabant are three ma-
a direct effect of rimonabant on peripheral tissues, such as
jor current weight loss medications (95). Orlistat is a gastric
the liver (101). Rimonabant has not been found to alter
and pancreatic lipase inhibitor (95). It reduces dietary fat
LDL-cholesterol or blood pressure consistently, and there
absorption by ⬃30% (96) and results in a mean weight loss
are currently no data on cardiovascular morbidity or mor-
of 3 kg (95). In a meta-analysis of 11 placebo-controlled
tality (95). However, several trials are underway, the largest
trials of 1 year, orlistat use has been shown to reduce blood
of which is the Comprehensive Rimonabant Evaluation
pressure, LDL-cholesterol, and fasting glucose in patients
Study of Cardiovascular Endpoints and Outcomes trial (95).
with diabetes (97,98). However, there are no long-term
This trial is studying the effect of rimonabant on myocardial
outcome data showing that orlistat use decreases major
infarction, stroke, and cardiovascular death in 17,000 obese
obesity-related morbidity and mortality (95).
patients (95).
Sibutramine is a combined serotonin and noradrenaline
reuptake inhibitor, which works primarily through its active
amine metabolites (99). Sibutramine produces a mean Efficacy of Exercise Compared with Dieting in Raising
weight loss of 4 to 5 kg (95), which is achieved predomi- HDL-cholesterol in Overweight Individuals
nantly by its centrally mediated action in increasing satiety, The two recommended non-pharmacologic approaches to
thereby reducing food intake (95,99). It also enhances en- raising plasma HDL-cholesterol in overweight individuals
ergy expenditure by stimulating thermogenesis—that is, it are exercise and diet. In two prospective studies, reductions
increases sympathetic activity to thermogenically active in energy intake to 1000 kcal/d produced modest weight
brown fat (95,99). This peripheral action of sibutramine, loss in patients of varying degrees of obesity (102,103). In
however, plays a minor part in weight reduction (95). Long- both studies, after sustained weight loss and a stable reduc-
term studies have not shown any significant effect of sib- tion in body weight, the total HDL-cholesterol levels in-
utramine on LDL-cholesterol and glycemic control (95). creased above baseline values. Similarly, in a study in
Similar to orlistat, there is a lack of long-term data on the severely obese individuals, both low-carbohydrate and con-
effect of sibutramine on major obesity-related morbidity ventional diets (with caloric restrictions imposed in both)
and mortality (95). There is, however, a current clinical trial produced similar (non-significant) increases in HDL-cho-
called the Sibutramine Cardiovascular Outcomes Trial, lesterol above baseline (104).
which is assessing the efficacy of sibutramine in reducing Although in one random, controlled, prospective study,
cardiac events such as myocardial infarction, stroke, and weight loss achieved through long-term calorie restriction
cardiovascular mortality in 9000 obese and overweight pa- was equally effective as exercise in raising total HDL-
tients (95). The Sibutramine Cardiovascular Outcomes Trial cholesterol levels in overweight individuals (105), dieting in
is expected to finish in 2008 (95). general has been shown to be less effective in altering HDL
Neither orlistat nor sibutramine produces a consistent levels than exercise. In obese male subjects, hypocaloric
increase in HDL-cholesterol levels (95). In contrast, rimon- dieting, in contrast to aerobic exercise (70), did not increase
abant, an endocannabinoid receptor antagonist, which re- the cardioprotective HDL2 and apo A-I subfractions and
duces weight by 4 to 5 kg, on average, and decreases waist actually diminished the effects of weight loss on increasing
2882 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
HDL2 levels (106). Also, weight loss achieved through the in HDL-cholesterol. Obviously, a randomly designed study
National Cholesterol Education Program’s recommended conducted on a large sample population group would tend to
low-saturated-fat, low-cholesterol diet did not alter HDL- increase the accuracy and probability of detecting signifi-
cholesterol levels in a 1-year random, controlled study of cant differences in HDL levels. Furthermore, a greater im-
164 men and women (107). Dietary fat, in fact, has been provement in HDL-cholesterol responses to exercise seems
shown to increase HDL levels in mice by increasing the to occur in men compared with women and in more severely
transport rates and decreasing the fractional catabolic rates obese individuals. Differences in baseline HDL-cholesterol
of HDL-cholesterol ester and apo A-I (108). levels among individuals may have contributed to the het-
erogeneous responses to exercise in different sample popu-
Importance of Experimental Design in Exercise lations. Subjects with higher initial HDL-cholesterol levels
Intervention Studies (women and individuals with less relative body weight and
Prospective studies to investigate whether increases in BMI) may not experience the magnitude of increase in
physical activity produce beneficial changes in plasma HDL HDL-cholesterol seen in subjects with lower baseline values
levels in overweight individuals have also produced con- (men and individuals with greater initial body weight and
flicting results. The response to exercise in these individuals BMI) (106,113).
has been found to be heterogeneous and dependent on the
following factors: 1) the methodologic approaches used; 2) Mechanisms of Exercise-induced Elevations in HDL
the intensity and duration of exercise; and 3) fitness levels Levels
achieved. Several studies have compared the effects of exercise and
In obese men and women, exercise programs of various weight loss per se (as a result of hypocaloric dieting) on
intensities and durations have been reported to increase
HDL-cholesterol changes in obese subjects. Wood et al.
(105,109), decrease (110), and not alter (111) HDL-choles-
(107) showed, in a random, 1-year controlled study of 84
terol levels. These variations in the reported effectiveness of
overweight sedentary men, that moderate exercise mediates
exercise on HDL-cholesterol have been attributed to differ-
a greater increase in total HDL- and HDL2-cholesterol per
ences in experimental design. For example, not all studies
kilogram body weight loss than hypocaloric dieting. Simi-
were controlled for collateral changes in diet, smoking, and
larly, Sopko et al. (118) reported that moderate exercise and
alcohol intake, all of which can substantially alter lipopro-
weight loss mediate separate and independent effects on
tein profiles (112,113).
HDL-cholesterol and that their effects are additive. The
Second, the timing of plasma collection can have a pro-
above studies, combined with the evidence that decreases in
found effect on HDL-cholesterol levels (112). Exercise ex-
erts an acute effect on plasma lipids, distinct from its body fat are more easily attained and maintained when
chronic effects because of training and weight loss. For exercise is part of a weight-loss program (119), underscore
example, 40 minutes of continuous treadmill exercise in the greater effectiveness of exercise in raising HDL-choles-
nine healthy women led to transient increases in HDL- terol levels than dieting.
cholesterol levels that returned to baseline values within 24 Exercise mediates a positive effect on HDL3b, HDL2b,
hours (114). Such increases in HDL with acute exercise and HDL apoA-I components, at least partly through
have been attributed to transient increases in lipoprotein changes in enzymes of lipoprotein metabolism. In one
lipase activity and clearance of triglyceride-rich lipoproteins study, the biological half-life of HDL apoA-I was found to
from the circulation (115), which, as discussed above, can be ⬃1.5-fold greater in endurance-trained men vs. sedentary
alter the composition and catabolic rate of HDLs. Thus, controls subjects (116). An 80% greater lipoprotein lipase
studies that measure the effects of exercise-induced weight activity and 38% lower hepatic lipase activity were also
loss on HDL-cholesterol levels at steady-state conditions found in the endurance-trained group. Therefore, increased
before, during, and after exercise sessions would tend to be lipid transfer to HDL by lipoprotein lipase and/or reduced
more reliable. HDL clearance by hepatic lipase were believed to account
Third, differences in plasma volumes across subjects for the prolonged apo A-I half-life in the trained men.
were not accounted for in all exercise intervention studies In a later prospective study, Kiens and Lithell (120)
(112). Because plasma volume expands with training, showed that endurance training directly increased lipopro-
plasma samples uncorrected for changes in an individual’s tein lipase activity in skeletal muscle tissue. Schwartz and
plasma volume would tend to underestimate the effect of Brunzell (121) found that, in obese individuals, lipoprotein
exercise on plasma HDL levels (116). Weight loss, con- lipase activity in adipose tissue increased with weight sta-
versely, reduces plasma volume, and, therefore, plasma bilization at a reduced weight (through hypocaloric dieting)
samples may overestimate its effect on HDL levels (117). and, conversely, returned to baseline levels with weight
Finally, the number and attributes of the subjects studied gain. Although it has not been specifically tested (to our
can contribute to differences in exercise-mediated changes knowledge), the independent effects of exercise and weight
OBESITY Vol. 15 No. 12 December 2007 2883
Obesity and High-density Lipoproteins, Rashid and Genest
loss per se may be attributed to different tissue-specific the fatty Zucker rat) should help in this regard. Also, more
alterations in lipoprotein lipase activity. studies on the effect of obesity on the molecular mecha-
nisms of HDL metabolism (receptor and post-receptor
Role of Exercise Intensity, Duration, and Training on mechanisms) need to be carried out.
HDL-cholesterol Finally, a caveat to consider in non-pharmacologic ap-
There is evidence to indicate that improvements in HDL- proaches to weight loss and increasing plasma HDL con-
cholesterol levels occur only when threshold exercise inten- centrations is the behavioral and metabolic resistance of
sities or duration are reached (122–126). Evaluation of overweight individuals to weight loss. Only 5% of over-
serum lipoproteins in overweight/obese postmenopausal weight subjects are able to achieve and maintain weight loss
women after 1 year of moderate-intensity exercise showed for a prolonged period of time (129). Metabolically, this
no change in HDL levels from baseline values (122). Wil- may partly be a result of an existing set point for fat mass or
liams et al. (124) reported that plasma HDL-cholesterol fat cell size (121). Therefore, a combination of behavioral,
concentrations in healthy, middle-age, sedentary men did diet, and exercise interventions may be required to attain
not change with endurance running unless they ran at least and maintain weight loss and improvements in HDL-cho-
10 mi/wk at 70% to 85% of maximum exercise intensity lesterol levels in obese subjects.
over a period of 9 months. Post hoc analysis of 6849
non-smoking runners of varying BMI further revealed that Acknowledgment
HDL-cholesterol levels increased by 0.2 mg/dL and BMI S.R. is supported by a Canadian Institutes of Health
decreased by 0.05 kg/m2 with every 1-mi increase in dis- Research Fellowship.
tance run per week (125). Further analysis revealed that the
number of miles run per week required to significantly References
increase HDL, decrease the total cholesterol-to-HDL ratio, 1. Gordon T, Castelli WP, Hjortland MC, Kannel WB,
and decrease triglycerides was much greater than the num- Dawber TR. High density lipoprotein as a protective factor
ber needed to reduce BMI. against coronary heart disease. The Framingham Study. Am J
Thompson et al. (127) observed that, even with prolonged Med. 1977;62:707–14.
exercise training (8 to 11 months) of high intensity (1 hour 2. von Eckardstein A, Huang Y, Assmann G. Physiological
role and clinical relevance of high-density lipoprotein sub-
supervised exercise training on bicycle ergometers at 80%
classes. Curr Opin Lipidol. 1994;5:404 –16.
of maximum heart rate), changes in HDL-cholesterol con- 3. Rader DJ. Molecular regulation of HDL metabolism and
centration are modest (an average of 13% or 5 mg/dL). function: implications for novel therapies. J Clin Invest.
However, because cardiovascular risk decreases by 1% to 2006;116:3090 –100.
2% for every 1- to 2-mg/dL increase in HDL (128) and 4. Ng DS, Leiter LA, Vezina C, Connelly PW, Hegele RA.
because physical activity per se is cardioprotective, weight Apolipoprotein A-I Q[-2]X causing isolated apolipoprotein
loss achieved through exercise likely represents substan- A-I deficiency in a family with analphalipoproteinemia.
tially decreased CAD risk in obese individuals. J Clin Invest. 1994;93:223–9.
5. Tilly-Kiesi M, Lichtenstein AH, Ordovas JM, et al. Sub-
jects with ApoA-I(Lys107–⬎0) exhibit enhanced fractional
catabolic rate of ApoA-I in Lp(AI) and ApoA-II in Lp(AI
Conclusions and Future Directions with AII). Arterioscler Thromb Vasc Biol. 1997;17:873– 80.
From the preceding sections, it is clear that excess weight 6. Chroni A, Duka A, Kan HY, Liu T, Zannis VI. Point
and adiposity are linked to low HDL-cholesterol levels in mutations in apolipoprotein A-I mimic the phenotype ob-
obese individuals. The fact that low HDL levels in obese served in patients with classical lecithin:cholesterol acyl-
states can be increased with reduction of adiposity suggests transferase deficiency. Biochemistry. 2005;44:14353– 66.
(but does not prove) a causal link. The relationship among 7. Mott S, Yu L, Marcil M, Boucher B, Rondeau C, Genest
obesity, HDL-cholesterol levels, and CAD risk could, per- J Jr. Decreased cellular cholesterol efflux is a common
haps, be more closely studied in humans by administering cause of familial hypoalphalipoproteinemia: role of the
an HDL-raising agent or placebo randomly to a select ABCA1 gene mutations. Atherosclerosis. 2000;152:457– 68.
population with low HDL-cholesterol levels and varying 8. Francis GA, Knopp RH, Oram JF. Defective removal of
degrees of obesity and examining clinical CAD endpoints. cellular cholesterol and phospholipids by apolipoprotein A-I
in Tangier Disease. J Clin Invest. 1995;96:78 – 87.
The effects of confounding factors affecting HDL-choles-
9. Albrecht C, Baynes K, Sardini A, et al. Two novel mis-
terol levels (e.g., smoking, alcohol, diet, other dyslipi- sense mutations in ABCA1 result in altered trafficking and
demias) would have to be carefully controlled. cause severe autosomal recessive HDL deficiency. Biochim
Furthermore, the mechanisms contributing to HDL-cho- Biophys Acta. 2004;1689:47–57.
lesterol lowering in obese subjects need to be more pre- 10. Emmerich J, Verges B, Tauveron I, et al. Familial HDL
cisely elucidated. Animal models that more closely mimic deficiency due to marked hypercatabolism of normal apoA-I.
the human lipoprotein profile than presently available (e.g., Arterioscler Thromb. 1993;13:1299 –306.
2884 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
11. Williams PT, Haskell WL, Vranizan KM, Krauss RM. level, and glucose intolerance as independent predictors of
The associations of high-density lipoprotein subclasses with the HDL2 cholesterol level in older adults. N Engl J Med.
insulin and glucose levels, physical activity, resting heart 1990;322:229 –34.
rate, and regional adiposity in men with coronary artery 26. Despres JP, Moorjani S, Ferland M, et al. Adipose tissue
disease: the Stanford Coronary Risk Intervention Project distribution and plasma lipoprotein levels in obese women.
baseline survey. Metab Clin Exp. 1995;44:106 –14. Importance of intra-abdominal fat. Arteriosclerosis. 1989;9:
12. Diehl AK, Fuller JH, Mattock MB, Salter AM, el Gohari 203–10.
R, Keen H. The relationship of high density lipoprotein 27. Kissebah AH, Vydelingum N, Murray R, et al. Relation of
subfractions to alcohol consumption, other lifestyle factors, body fat distribution to metabolic complications of obesity.
and coronary heart disease. Atherosclerosis. 1988;69: J Clin Endocrinol Metab. 1982;54:254 – 60.
145–53. 28. Nicklas BJ, Penninx BW, Ryan AS, Berman DM, Lynch
13. von Eckardstein A, Assmann G. Prevention of coronary NA, Dennis KE. Visceral adipose tissue cutoffs associated
heart disease by raising high-density lipoprotein cholesterol? with metabolic risk factors for coronary heart disease in
Curr Opin Lipidol. 2001;11:627–37. women. Diabetes Care. 2003;26:1413–20.
14. Robins SJ. Targeting low high-density lipoprotein choles- 29. Despres JP, Couillard C, Gagnon J, et al. Race, visceral
terol for therapy: lessons from the Veterans Affairs High- adipose tissue, plasma lipids, and lipoprotein lipase activity
density Lipoprotein Intervention Trial. Am J Cardiol. 2001; in men and women: the Health, Risk Factors, Exercise Train-
88:19N–23N. ing, and Genetics (HERITAGE) family study. Arterioscler
15. Despres JP, Moorjani S, Lupien PJ, Tremblay A, Nadeau Thromb Vasc Biol. 2000;20:1932– 8.
A, Bouchard C. Regional distribution of body fat, plasma 30. Lear SA, Toma M, Birmingham CL, Frohlich JJ. Modi-
lipoproteins, and cardiovascular disease. Arterioscler. 1990; fication of the relationship between simple anthropometric
10:497–511. indices and risk factors by ethnic background. Metabolism.
16. Campfield LA, Smith FJ, Burn P. Strategies and potential 2003;52:1295–301.
molecular targets for obesity treatment. Science. 1998;280: 31. Schmitz G, Lackner KJ. High-density lipoproteins and
1383–7. atherosclerosis. Curr Opin Lipidol. 1993;4:392– 400.
17. Hubert HB, Feinleib M, McNamara PM, Castelli WP. 32. Dobiasova M, Frohlich J. Understanding the mechanism of
Obesity as an independent risk factor for cardiovascular LCAT reaction may help to explain the high predictive value
disease: a 26-year follow-up of participants in the Framing- of LDL/HDL-cholesterol ratio. Physiol Res. 1998;47:
ham Heart Study. Circulation. 1983;67:968 –77. 387–97.
18. Killeen J, Vanderburg D, Harlan WR. Application of weight- 33. Bertiere MC, Fumeron F, Rigaud D, Malon D, Apfel-
height ratios and body indices to juvenile populations—the baum M, Girard-Globa A. Low high density lipoprotein-2
National Health Examination Survey Data. J Chronic Dis. concentrations in obese male subjects. Atherosclerosis. 1988;
1978;31:529 –37. 73:57– 61.
19. Toth PP. High-density lipoprotein as a therapeutic target: 34. Carr MC, Hokanson JE, Zambon A, et al. The contribu-
clinical evidence and treatment strategies. Am J Cardiol. tion of intra-abdominal fat to gender differences in hepatic
2005;96:50K– 8K. lipase activity and low/high density lipoprotein heterogene-
20. Glueck CJ, Taylor HL, Jacobs D, Morrison JA, Beagle- ity. J Clin Endocrinol Metab. 2001;86:2831–7.
hole R, Williams OD. Plasma high density lipoprotein 35. Miller NE. Associations of high-density lipoprotein sub-
cholesterol: association with measurements of body mass. classes and apolipoproteins with ischemic heart disease and
Circulation. 1980;62(Suppl IV):IV62–9. coronary atherosclerosis. Am Heart J. 1987;113:589 –97.
21. Lamon-Fava S, Wilson PW, Schaefer EJ. Impact of body 36. Patsch JR, Prasad S, Gotto AM Jr, Bengtsson-Olivecrona
mass index on coronary heart disease risk factors in men and G. Postprandial lipemia. A key for the conversion of high
women. The Framingham Offspring Study. Arterioscler density lipoprotein2 into high density lipoprotein3 by hepatic
Thromb Vasc Biol. 1996;16:1509 –15. lipase. J Clin Invest. 1984;74:2017–23.
22. Larsson B, Svardsudd K, Welin L, Wilhelmsen L, Björn- 37. Despres JP, Ferland M, Moorjani S, et al. Role of hepatic-
torp P, Tibblin G. Abdominal adipose tissue distribution, triglyceride lipase activity in the association between intra-
obesity, and risk of cardiovascular disease and death: 13 year abdominal fat and plasma HDL-cholesterol in obese women.
follow up of participants in the study of men born in 1913. Arteriosclerosis. 1989;9:485–92.
Br Med J (Clin Res Ed). 1984;288:1401– 4. 38. Patsch JR, Karlin JB, Scott LW, Smith LC, Gotto AM Jr.
23. Peiris AN, Sothmann MS, Hoffmann RG, et al. Adiposity, Inverse relationship between blood levels of high density
fat distribution, and cardiovascular risk. Ann Intern Med. 1989; lipoprotein subfraction 2 and magnitude of postprandial
110:867–72. lipemia. Proc Natl Acad Sci USA. 1983;80:1449 –53.
24. Kissebah AH, Alfarsi S, Adams PW. Integrated regu- 39. Tall AR. Plasma high density lipoproteins. Metabolism and
lation of very low density lipoprotein triglyceride and relationship to atherogenesis. J Clin Invest. 1990;86:379 – 84.
apolipoprotein-B kinetics in man: normolipemic subjects, 40. Ginsberg HN. Insulin resistance and cardiovascular disease.
familial hypertriglyceridemia and familial combined hyper- J Clin Invest. 2000;106:453– 8.
lipidemia. Metab Clin Exp. 1981;30:856 – 68. 41. Coppack SW, Evans RD, Fisher RM, et al. Adipose tissue
25. Ostlund RE Jr, Staten M, Kohrt WM, Schultz J, Malley metabolism in obesity: lipase action in vivo before and after
M. The ratio of waist-to-hip circumference, plasma insulin a mixed meal. Metab Clin Exp. 1992;41:264 –72.
OBESITY Vol. 15 No. 12 December 2007 2885
Obesity and High-density Lipoproteins, Rashid and Genest
42. Haemmerle G, Zimmermann R, Zechner R. Letting lipids 56. Jahangiri A, Rader DJ, Marchadier D, Curtiss LK, Bon-
go: hormone-sensitive lipase. Curr Opin Lipidol. 2003;14: net DJ, Rye KA. Evidence that endothelial lipase remodels
289 –97. high density lipoproteins without mediating the dissociation
43. Murakami T, Michelagnoli S, Longhi R, et al. Triglycer- of apolipoprotein A-I. J Lipid Res. 2005;46:896 –903.
ides are major determinants of cholesterol esterification/ 57. Paradis ME, Badellino KO, Rader DJ, et al. Visceral
transfer and HDL remodeling in human plasma. Arterioscler adiposity and endothelial lipase. J Clin Endocrinol Metab.
Thromb Vasc Biol. 1995;15:1819 –28. 2006;91:3538 – 43.
44. Arai T, Yamashita S, Hirano K, et al. Increased plasma 58. Zimmermann R, Strauss JG, Haemmerle G, et al. Fat
cholesteryl ester transfer protein in obese subjects. A mobilization in adipose tissue is promoted by adipose tri-
possible mechanism for the reduction of serum HDL- glyceride lipase. Science. 2004;306:1383– 6.
cholesterol levels in obesity. Arterioscler Thromb. 1994; 59. Schweiger M, Schreiber R, Haemmerle G, et al. Adipose
14:1129 –36. triglyceride lipase and hormone-sensitive lipase are the major
45. McLaughlin T, Abbasi F, Lamendola C, Reaven G. Het-
enzymes in adipose tissue triacylglycerol catabolism. J Biol
erogeneity in the prevalence of risk factors for cardiovascular
Chem. 2006;281:40236 – 41.
disease and type 2 diabetes mellitus in obese individuals:
60. Langin D, Dicker A, Tavernier G, et al. Adipocyte lipases
effect of differences in insulin sensitivity. Arch Intern Med.
and defect of lipolysis in human obesity. Diabetes. 2005;54:
2007;167:642– 8.
3190 –7.
46. Sorbris R, Petersson BG, Nilsson-Ehle P. Effects of
weight reduction on plasma lipoproteins and adipose tis- 61. Jocken JW, Langin D, Smit E, et al. Adipose triglyceride
sue metabolism in obese subjects. Eur J Clin Invest. lipase and hormone-sensitive lipase protein expression is
1981;11:491– 8. decreased in the obese insulin-resistant state. J Clin Endo-
47. McKenney JM, Davidson MH, Shear CL, Revkin JH. crinol Metab. 2007;92:2292–9.
Efficacy and safety of torcetrapib, a novel cholesteryl 62. Jauhiainen M, Metso J, Pahlman R, Blomqvist S, van Tol
ester transfer protein inhibitor, in individuals with below- A, Ehnholm C. Human plasma phospholipid transfer protein
average high-density lipoprotein cholesterol levels on a causes high density lipoprotein conversion. J Biol Chem.
background of atorvastatin. J Am Coll Cardiol. 2006;48: 1993;268:4032– 6.
1782–90. 63. Marques-Vidal P, Jauhiainen M, Metso J, Ehnholm C.
48. Clark RW, Ruggeri RB, Cunningham D, Bamberger MJ. Transformation of high density lipoprotein 2 particles by he-
Description of the torcetrapib series of cholesteryl ester patic lipase and phospholipid transfer protein. Atherosclerosis.
transfer protein inhibitors, including mechanism of action. 1997;133:87–95.
J Lipid Res. 2006;47:537–52. 64. Dullaart RP, Sluiter WJ, Dikkeschei LD, Hoogenberg K,
49. Carr MC, Ayyobi AF, Murdoch SJ, Deeb SS, Brunzell van Tol A. Effect of adiposity on plasma lipid transfer
JD. Contribution of hepatic lipase, lipoprotein lipase, and protein activities: a possible link between insulin resistance
cholesteryl ester transfer protein to LDL and HDL heteroge- and high density lipoprotein metabolism. Eur J Clin Invest.
neity in healthy women. Arterioscler Thromb Vasc Biol. 1994;24:188 –94.
2002;22:667–73. 65. Kaser S, Laimer M, Sandhofer A, Salzmann K, Eben-
50. Foger B, Ritsch A, Doblinger A, Wessels H, Patsch JR. bichler CF, Patsch JR. Effects of weight loss on PLTP
Relationship of plasma cholesteryl ester transfer protein to activity and HDL particle size. Int J Obes Relat Metab
HDL-cholesterol. Studies in normotriglyceridemia and mod- Disord. 2004;28:1280 –2.
erate hypertriglyceridemia. Arterioscler Thromb Vasc Biol. 66. Williams PT, Wood PD, Krauss RM, et al. Does weight
1996;16:1430 – 6. loss cause the exercise-induced increase in plasma high den-
51. Borggreve SE, De Vries R, Dullaart RP. Alterations in high-
sity lipoproteins? Atherosclerosis. 1983;47:173– 85.
density lipoprotein metabolism and reverse cholesterol transport
67. Wolf RN, Grundy SM. Effects of caloric restriction on
in insulin resistance and type 2 diabetes mellitus: role of lipo-
plasma lipids and lipoproteins. Clin Res. 1980;28:55a.
lytic enzymes, lecithin:cholesterol acyltransferase and lipid
68. Fong BS, Rodrigues PO, Salter AM, et al. Characterization
transfer proteins. Eur J Clin Invest. 2003;33:1051– 69.
of high density lipoprotein binding to human adipocyte
52. Berenson A. End of trial is a big loss for Pfizer and heart
patients. New York Times. 2006;4:A26. plasma membranes. J Clin Invest. 1985;75:1804 –12.
53. Hamilton JA, Deckelbaum RJ. Crystal structure of CETP: 69. Despres JP, Fong BS, Julien P, Jimenez J, Angel A.
new hopes for raising HDL to decrease risk of cardiovascular Regional variation in HDL metabolism in human fat cells:
disease? Nat Struct Mol Biol. 2007;14:95–7. effect of cell size. Am J Physiol. 1987;252:E654 –9.
54. Howes LG, Kostner K. The withdrawal of torcetrapib from 70. Schwartz RS. The independent effects of dietary weight loss
drug development: implications for the future of drugs that and aerobic training on high density lipoproteins and apo-
alter HDL metabolism. Expert Opin Investig Drugs. 2007; lipoprotein A-I concentrations in obese men. Metabolism.
16:1509 –16. 1987;36:165–71.
55. Qiu X, Mistry A, Ammirati MJ, et al. Crystal structure of 71. Maciejko JJ, Holmes DR, Kottke BA, Zinsmeister AR,
cholesteryl ester transfer protein reveals a long tunnel and Dinh DM, Mao SJ. Apolipoprotein A-I as a marker of
four bound lipid molecules. Nat Struct Mol Biol. 2007;14: angiographically assessed coronary-artery disease. N Engl
106 –13. J Med. 1983;309:385–9.
2886 OBESITY Vol. 15 No. 12 December 2007
Obesity and High-density Lipoproteins, Rashid and Genest
72. Kunitake ST, La Sala KJ, Kane JP. Apolipoprotein A-I- 87. Sasahara T, Yamashita T, Sviridov D, Fidge N, Nestel P.
containing lipoproteins with pre-beta electrophoretic mobil- Altered properties of high density lipoprotein subfractions in
ity. J Lipid Res. 1985;26:549 –55. obese subjects. J Lipid Res. 1997;38:600 –11.
73. Garrison RJ, Wilson PW, Castelli WP, Feinleib M, Kan- 88. Krimbou L, Marcil M, Genest J. New insights into the
nel WB, McNamara PM. Obesity and lipoprotein choles- biogenesis of human high-density lipoproteins. Curr Opin
terol in the Framingham offspring study. Metabolism. 1980; Lipidol. 2006;17:258 – 67.
29:1053– 60. 89. Krimbou L, Hajj HH, Blain S, et al. Biogenesis and
74. Oram JF. Effects of high density lipoprotein subfractions on speciation of nascent apoA-I-containing particles in various
cholesterol homeostasis in human fibroblasts and arterial cell lines. J Lipid Res. 2005;46:1668 –77.
smooth muscle cells. Arteriosclerosis. 1983;3:420 –32. 90. Francone OL, Gurakar A, Fielding C. Distribution and
75. Khoo JC, Miller E, McLoughlin P, Steinberg D. Preven- functions of lecithin:cholesterol acyltransferase and cho-
tion of low density lipoprotein aggregation by high density lesteryl ester transfer protein in plasma lipoproteins. Evi-
lipoprotein or apolipoprotein A-I. J Lipid Res. 1990;31:645– dence for a functional unit containing these activities to-
52. gether with apolipoproteins A-I and D that catalyzes the
76. Rubin EM, Krauss RM, Spangler EA, Verstuyft JG, Clift esterification and transfer of cell-derived cholesterol. J Biol
SM. Inhibition of early atherogenesis in transgenic mice by Chem. 1989;264:7066 –72.
human apolipoprotein AI. Nature. 1991;353:265–7. 91. Huang Y, von Eckardstein A, Assmann G. Cell-derived
77. Brinton EA, Eisenberg S, Breslow JL. Increased apo A-I unesterified cholesterol cycles between different HDLs and
and apo A-II fractional catabolic rate in patients with low LDL for its effective esterification in plasma. Arterioscler
high density lipoprotein-cholesterol levels with or without Thromb. 1993;13:445–58.
hypertriglyceridemia. J Clin Invest. 1991;87:536 – 44. 92. Kaser S, Sandhofer A, Foger B, et al. Influence of obesity
78. Gylling H, Vega GL, Grundy SM. Physiologic mechanisms and insulin sensitivity on phospholipid transfer protein ac-
for reduced apolipoprotein A-I concentrations associated tivity. Diabetologia. 2001;44:1111–7.
with low levels of high density lipoprotein cholesterol in 93. Sasahara T, Nestel P, Fidge N, Sviridov D. Cholesterol
patients with normal plasma lipids. J Lipid Res. 1992;33: transport between cells and high density lipoprotein subfrac-
1527–39. tions from obese and lean subjects. J Lipid Res. 1998;39:
79. Ooi EM, Watts GF, Farvid MS, et al. High-density li- 544 –54.
poprotein apolipoprotein A-I kinetics in obesity. Obes Res. 94. Miida T, Nakamura Y, Inano K, et al. Pre beta 1-high-
2005;13:1008 –16. density lipoprotein increases in coronary artery disease. Clin
80. Brinton EA, Eisenberg S, Breslow JL. Elevated high den- Chem. 1996;42:1992–5.
sity lipoprotein cholesterol levels correlate with decreased 95. Padwal RS, Majumdar SR. Drug treatments for obesity:
apolipoprotein A-I and A-II fractional catabolic rate in orlistat, sibutramine, and rimonabant. Lancet. 2007;369:
women. J Clin Invest. 1989;84:262–9. 71–7.
81. Brinton EA, Eisenberg S, Breslow JL. A low-fat diet 96. Hauptman JB, Jeunet FS, Hartmann D. Initial studies in
decreases high density lipoprotein (HDL) cholesterol levels humans with the novel gastrointestinal lipase inhibitor Ro
by decreasing HDL apolipoprotein transport rates. J Clin 18 – 0647 (tetrahydrolipstatin). Am J Clin Nutr. 1992;55
Invest. 1990;85:144 –51. (1 Suppl):309S–13S.
82. Brinton EA, Terry JG. Increased intra-abdominal fat may 97. Padwal R, Li SK, Lau DC. Long-term pharmacotherapy for
lower HDL levels by increasing the fractional catabolic rate obesity and overweight. Cochrane Database Syst Rev. 2004;
of lp A-I in postmenopausal women. Circulation. 1996;94 4:CD004094.
(Suppl):I-266. 98. Padwal R, Li SK, Lau DC. Long-term pharmacotherapy for
83. Dullaart RP, Sluiter WJ, Dikkeschei LD, Hoogenberg K, overweight and obesity: a systematic review and meta-
Van Tol A. Effect of adiposity on plasma lipid transfer analysis of randomized controlled trials. Int J Obes Relat
protein activities: a possible link between insulin resistance Metab Disord. 2003;27:1437– 46.
and high density lipoprotein metabolism. Eur J Clin Invest. 99. Nisoli E, Carruba MO. An assessment of the safety and
1994;24:188 –94. efficacy of sibutramine, an anti-obesity drug with a novel
84. Deeb SS, Zambon A, Carr MC, Ayyobi AF, Brunzell JD. mechanism of action. Obes Rev. 2000;1:127–39.
Hepatic lipase and dyslipidemia: interactions among genetic 100. Pi-Sunyer FX, Aronne LJ, Heshmati HM, Devin J,
variants, obesity, gender, and diet. J Lipid Res. 2003;44: Rosenstock J. Effect of rimonabant, a cannabinoid-1 recep-
1279 – 86. tor blocker, on weight and cardiometabolic risk factors in
85. Rashid S, Barrett PH, Uffelman KD, Watanabe T, Adeli overweight or obese patients: RIO-North America: a ran-
K, Lewis GF. Lipolytically modified triglyceride-enriched domized controlled trial. JAMA. 2006;295:761–75.
HDLs are rapidly cleared from the circulation. Arterioscler 101. Henness S, Robinson DM, Lyseng-Williamson KA.
Thromb Vasc Biol. 2002;22:483–7. Rimonabant. Drugs. 2006;66:2109 –19.
86. Rashid S, Uffelman K, Lewis GF. The mechanism of 102. Wolf RN, Grundy SM. Influence of weight reduction on
HDL lowering in hypertriglyceridemic, insulin-resistant plasma lipoproteins in obese patients. Arteriosclerosis. 1983;
states. J Diabetes Complications. 2002;16:24 – 8. 3:160 –9.
OBESITY Vol. 15 No. 12 December 2007 2887
Obesity and High-density Lipoproteins, Rashid and Genest
103. Rossner S, Bjorvell H. Early and late effects of weight loss Acute increase in lipoprotein lipase following prolonged
on lipoprotein metabolism in severe obesity. Atherosclerosis. exercise. Metabolism. 1984;33:454 –7.
1987;64:125–30. 116. Herbert PN, Bernier DN, Cullinane EM, Edelstein L,
104. Seshadri P, Iqbal N, Stern L, et al. A randomized study Kantor MA, Thompson PD. High-density lipoprotein me-
comparing the effects of a low-carbohydrate diet and a con- tabolism in runners and sedentary men. JAMA. 1984;252:
ventional diet on lipoprotein subfractions and C-reactive 1034 –7.
protein levels in patients with severe obesity. Am J Med. 117. Reisin E, Frohlich ED, Messerli FH, et al. Cardiovascular
2004;117:398 – 405. changes after weight reduction in obesity hypertension. Ann
105. Wood PD, Stefanick ML, Dreon DM, et al. Changes in Intern Med. 1983;98:315–9.
plasma lipids and lipoproteins in overweight men during 118. Sopko G, Leon AS, Jacobs DR Jr, et al. The effects of
weight loss through dieting as compared with exercise. exercise and weight loss on plasma lipids in young obese
N Engl J Med. 1988;319:1173–9. men. Metabolism. 1985;34:227–36.
106. Williams PT, Krauss RM, Vranizan KM, Wood PD. 119. Martin JE, Dubbert PM. Exercise applications and promo-
Changes in lipoprotein subfractions during diet-induced and tion in behavioral medicine: current status and future direc-
exercise-induced weight loss in moderately overweight men. tions. J Consult Clin Psychol. 1982;50:1004 –17.
Circulation. 1990;81:1293–304. 120. Kiens B, Lithell H. Lipoprotein metabolism influenced by
107. Wood PD, Stefanick ML, Williams PT, Haskell WL. The training-induced changes in human skeletal muscle. J Clin
effects on plasma lipoproteins of a prudent weight-reducing Invest. 1989;83:558 – 64.
diet, with or without exercise, in overweight men and 121. Schwartz RS, Brunzell JD. Increase of adipose tissue li-
women. N Engl J Med. 1991;325:461– 6.
poprotein lipase activity with weight loss. J Clin Invest.
108. Hayek T, Ito Y, Azrolan N, et al. Dietary fat increases high
1981;67:1425–30.
density lipoprotein (HDL) levels both by increasing the
122. Mohanka M, Irwin M, Heckbert SR, et al. Serum li-
transport rates and decreasing the fractional catabolic rates of
poproteins in overweight/obese postmenopausal women: a
HDL-cholesterol ester and apolipoprotein (Apo) A-I. Presen-
one-year exercise trial. Med Sci Sports Exerc. 2006;38:
tation of a new animal model and mechanistic studies in
231–9.
human Apo A-I transgenic and control mice. J Clin Invest.
123. Park SK, Park JH, Kwon YC, Kim HS, Yoon MS, Park
1993;91:1665–71.
HT. The effect of combined aerobic and resistance exercise
109. Williams PT. Lipoproteins and adiposity show improvement
training on abdominal fat in obese middle-aged women.
at substantially higher exercise levels than those currently
recommended. American Heart Association Conference 66th J Physiol Anthropol Appl Human Sci. 2003;22:129 –35.
Scientific Sessions. Atlanta, GA; November 8 –11, 1993. 124. Williams PT, Wood PD, Haskell WL, Vranizan K. The
110. Leon AS, Conrad J, Hunninghake DB, Serfass R. Effects effects of running mileage and duration on plasma lipopro-
of a vigorous walking program on body composition, and tein levels. JAMA. 1982;247:2674 –9.
carbohydrate and lipid metabolism of obese young men. 125. Williams PT. Weight set-point theory and the high-density
Am J Clin Nutr. 1979;32:1776 – 87. lipoprotein concentrations of long-distance runners. Metab-
111. Lewis S, Haskell WL, Wood PD, Manoogian N, Bailey olism. 1990;39:460 –7.
JE, Pereira MB. Effects of physical activity on weight 126. Thompson PD, Yurgalevitch SM, Flynn MM, et al. Effect
reduction in obese middle-aged women. Am J Clin Nutr. of prolonged exercise training without weight loss on high-
1976;29:151– 6. density lipoprotein metabolism in overweight men. Metabo-
112. Thompson PD. What do muscles have to do with lipopro- lism. 1997;46:217–23.
teins? Circulation. 1990;81:1428 –30. 127. Thompson PD, Cullinane EM, Sady SP, et al. Modest
113. Brownell KD, Bachorik PS, Ayerle RS. Changes in plasma changes in high-density lipoprotein concentration and me-
lipid and lipoprotein levels in men and women after a pro- tabolism with prolonged exercise training. Circulation. 1988;
gram of moderate exercise. Circulation. 1982;65:477– 84. 78:25–34.
114. Swank AM, Robertson RJ, Deitrich RW, Bates M. The 128. Rifkind BM. Gemfibrozil, lipids, and coronary risk. N Engl
effect of acute exercise on high density lipoprotein- J Med. 1987;317:1279 – 81.
cholesterol and the subfractions in females. Atherosclerosis. 129. Wing RR, Jeffery RW. Outpatient treatments of obesity: a
1987;63:187–92. comparison of methodology and clinical results. Int J Obes
115. Kantor MA, Cullinane EM, Herbert PN, Thompson PD. Relat Metab Disord. 1979;3:261–79.
2888 OBESITY Vol. 15 No. 12 December 2007
View publication stats
You might also like
- The HDL Handbook: Biological Functions and Clinical ImplicationsFrom EverandThe HDL Handbook: Biological Functions and Clinical ImplicationsNo ratings yet
- Srikant H 2016Document10 pagesSrikant H 2016aditya sekarNo ratings yet
- Elevated-LDL-cholesterol-levels-among-lean-mass-hyDocument4 pagesElevated-LDL-cholesterol-levels-among-lean-mass-hy8v6y5jsykbNo ratings yet
- HDL in CKD-ThesisDocument27 pagesHDL in CKD-ThesisEndy SusenoNo ratings yet
- To Eat or Not To Eat Dietary Fat in Uremia Is The QuestionDocument6 pagesTo Eat or Not To Eat Dietary Fat in Uremia Is The QuestionUriel FurlongNo ratings yet
- Effect of haemodialysis on serum lipids and biochemical markers in CKD patientsDocument4 pagesEffect of haemodialysis on serum lipids and biochemical markers in CKD patientsmelda lestari haidirNo ratings yet
- Raising HDL Cholesterol in Women - 111709 PDFDocument11 pagesRaising HDL Cholesterol in Women - 111709 PDFDina Aulia FakhrinaNo ratings yet
- Effect of Low Carbohydrate High Fat Diet On LDL Cholesterol and Gene Expression in Normal Weight, Young Adults A Randomized Controlled StudyDocument10 pagesEffect of Low Carbohydrate High Fat Diet On LDL Cholesterol and Gene Expression in Normal Weight, Young Adults A Randomized Controlled StudyJennie ManobanNo ratings yet
- GrasasDocument12 pagesGrasasCristian Camilo SaavedraNo ratings yet
- Journal Pre-ProofDocument28 pagesJournal Pre-Proofhenry hernandezNo ratings yet
- Ijms 24 16504Document13 pagesIjms 24 16504orthofitness2017No ratings yet
- Triglyceride Profile in Dyslipidaemia of Type 2 Diabetes MellitusDocument4 pagesTriglyceride Profile in Dyslipidaemia of Type 2 Diabetes MellitusTanveerNo ratings yet
- Metabolic SyndromeDocument7 pagesMetabolic SyndromeDannop GonzálezNo ratings yet
- Relationship between Dyslipidemia and Glycemic Status in Type-2 Diabetes MellitusDocument5 pagesRelationship between Dyslipidemia and Glycemic Status in Type-2 Diabetes Mellitusoliffasalma atthahirohNo ratings yet
- ProjectDocument32 pagesProjectNithya sudevNo ratings yet
- Management of HyperlipidemiaDocument34 pagesManagement of HyperlipidemiaCarleta StanNo ratings yet
- A Review On Causes and Risk Factors of HyperlipidemiaDocument6 pagesA Review On Causes and Risk Factors of HyperlipidemiaHam jungNo ratings yet
- Pattern of Dyslipidemia in Type Diabetic Subjects in Eastern NepalDocument3 pagesPattern of Dyslipidemia in Type Diabetic Subjects in Eastern NepalDR VENKATARAMANA MurthyNo ratings yet
- Risk Factors for Heart Disease Among Female University StudentsDocument6 pagesRisk Factors for Heart Disease Among Female University StudentsMuhammad Irfan JamilNo ratings yet
- 1.HDL-a Therapeutic TargetDocument8 pages1.HDL-a Therapeutic TargetAshraf ChowdhuryNo ratings yet
- Paradox of Hypercholesterolaemia in Highly Trained, Keto-Adapted AthletesDocument10 pagesParadox of Hypercholesterolaemia in Highly Trained, Keto-Adapted AthletesTennis workbookNo ratings yet
- Dyslipidemia in Diabetes Mellitus and Cardiovascular DiseaseDocument6 pagesDyslipidemia in Diabetes Mellitus and Cardiovascular DiseaseEward Rod SalNo ratings yet
- Functional Food and Bioactive Compounds On The Modulation of The Functionality of HDL-CDocument30 pagesFunctional Food and Bioactive Compounds On The Modulation of The Functionality of HDL-CSANDRA FERNANDES - NutriçãoNo ratings yet
- Significance of Orlistat for Dyslipidemia, SBP, and BMIDocument7 pagesSignificance of Orlistat for Dyslipidemia, SBP, and BMIGestne AureNo ratings yet
- Clase 1. Lipoproteins and Fatty Acids in CKD Molecular and Metabolic AlterationsDocument15 pagesClase 1. Lipoproteins and Fatty Acids in CKD Molecular and Metabolic Alterationsdulce orozcoNo ratings yet
- Journal Article-Advances - in - Treatment - of - DyslipidemiaDocument19 pagesJournal Article-Advances - in - Treatment - of - DyslipidemiaLauv LoveNo ratings yet
- Lipids and Lipoproteins in Patients With Type 2 Diabetes: Reviews/Commentaries/PositionstatementsDocument9 pagesLipids and Lipoproteins in Patients With Type 2 Diabetes: Reviews/Commentaries/PositionstatementsM Fadhil Wasi PradiptaNo ratings yet
- Endocrinology and Metabolism Clinics-Lípidos 2009Document234 pagesEndocrinology and Metabolism Clinics-Lípidos 2009Tony Miguel Saba SabaNo ratings yet
- Volek 2009Document13 pagesVolek 2009Paul SimononNo ratings yet
- Paradox of Hypercholesterolaemia in Highly Trained, Keto-Adapted AthletesDocument10 pagesParadox of Hypercholesterolaemia in Highly Trained, Keto-Adapted AthletesYo Vivo Fit Pablo y KarlaNo ratings yet
- Omega 3Document8 pagesOmega 3Mireya AlejandraNo ratings yet
- Dyslipidemia Management in Women and Men: Exploring Potential Gender DifferencesDocument7 pagesDyslipidemia Management in Women and Men: Exploring Potential Gender DifferencesAmelie AnsonNo ratings yet
- Review of Hyperlipidemia Types, Treatments, and New Drug TargetsDocument12 pagesReview of Hyperlipidemia Types, Treatments, and New Drug TargetsPNo ratings yet
- The Anti-Inflammatory Function of High-Density Lipoprotein in Type II Diabetes A Systematic ReviewDocument18 pagesThe Anti-Inflammatory Function of High-Density Lipoprotein in Type II Diabetes A Systematic ReviewA'in Qurrota A'yuninNo ratings yet
- HDL's Role in Reducing Atherosclerosis Risk via Its Antioxidant EffectsDocument7 pagesHDL's Role in Reducing Atherosclerosis Risk via Its Antioxidant EffectsDainius JakubauskasNo ratings yet
- KK 2Document4 pagesKK 2Arsal Wildan wildanNo ratings yet
- Effects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionDocument14 pagesEffects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionYoungFanjiensNo ratings yet
- Effect of Bariatric Surgery-Induced Weight Loss On SR-BI-, ABCG1-, and ABCA1-Mediated Cellular Cholesterol Efflux in Obese WomenDocument9 pagesEffect of Bariatric Surgery-Induced Weight Loss On SR-BI-, ABCG1-, and ABCA1-Mediated Cellular Cholesterol Efflux in Obese Womensergiuosan96No ratings yet
- A Hyper - Emia A A PseudohyponatremiaDocument14 pagesA Hyper - Emia A A PseudohyponatremiaSrinivas PolikepatiNo ratings yet
- Quizlet AntihyperlipidDocument4 pagesQuizlet AntihyperlipidLisetteNo ratings yet
- 7.systematic Review and Meta-Analysis of Clinical Trials of The Effects of Low Carbohydrate Diets On Cardiovascular Risk Factors.Document19 pages7.systematic Review and Meta-Analysis of Clinical Trials of The Effects of Low Carbohydrate Diets On Cardiovascular Risk Factors.Ambar Sofía MieresNo ratings yet
- Faktor Risiko PJK - Shabana - 2020Document7 pagesFaktor Risiko PJK - Shabana - 2020Ikhsanuddin QothiNo ratings yet
- Secondary Causes of DyslipidemiaDocument3 pagesSecondary Causes of DyslipidemiaAnghelo Aldair Velásquez CarrilloNo ratings yet
- Mty1107 Sec9 Saquiton MF LipidsfornursingDocument8 pagesMty1107 Sec9 Saquiton MF LipidsfornursingCes SaquitonNo ratings yet
- 8731 Ch02Document51 pages8731 Ch02Mohammed Salim AmmorNo ratings yet
- Case For Not Restricting Saturated Fat On Low Carb Diet-Nutrition Metab 2005 2 21-Volek ForsytheDocument2 pagesCase For Not Restricting Saturated Fat On Low Carb Diet-Nutrition Metab 2005 2 21-Volek Forsythedonmd98No ratings yet
- Phytosterols in The Treatment of Hypercholesterolemia and Prevention of Cardiovascular DiseasesDocument8 pagesPhytosterols in The Treatment of Hypercholesterolemia and Prevention of Cardiovascular DiseasesJherson Gustavo SánchezNo ratings yet
- Raza2004 PDFDocument12 pagesRaza2004 PDFIvana RuseskaNo ratings yet
- 6340 25219 1 PB PDFDocument3 pages6340 25219 1 PB PDFChris Jo ManojNo ratings yet
- Comparative Study of Atorvastatin and RosuvastatinDocument8 pagesComparative Study of Atorvastatin and RosuvastatinDewi nur thohidahNo ratings yet
- 13 FullDocument16 pages13 FullGyse LemaNo ratings yet
- Bjbabs 22 137Document10 pagesBjbabs 22 137Bilal BilimNo ratings yet
- ObeseDocument8 pagesObesesaravana kumarNo ratings yet
- Lawrence 2013 - Dietary Fats and Health - Dietary Recommendations in The Context of Scientific EvidenceDocument9 pagesLawrence 2013 - Dietary Fats and Health - Dietary Recommendations in The Context of Scientific EvidenceAlbert CalvetNo ratings yet
- pownall2021Document12 pagespownall2021AldoNo ratings yet
- Efficacy and Safety of High-Density Lipoprotein Cholesterol-Increasing CompoundsDocument13 pagesEfficacy and Safety of High-Density Lipoprotein Cholesterol-Increasing CompoundsAdityasekar WangiNo ratings yet
- Practical Laboratory Medicine: Subashini C. Thambiah, Leslie Charles LaiDocument21 pagesPractical Laboratory Medicine: Subashini C. Thambiah, Leslie Charles LaiAdina BrahaNo ratings yet
- Dislipidemia Dan CardiovasculerDocument2 pagesDislipidemia Dan CardiovasculerWahyuningsih HamidNo ratings yet
- Should Glycemic Index and Glycemic Load Be Considered in Dietary RecommendationsDocument22 pagesShould Glycemic Index and Glycemic Load Be Considered in Dietary RecommendationsLilo LiliNo ratings yet
- HDL Muy Alto 2019Document10 pagesHDL Muy Alto 2019Luis C Ribon VNo ratings yet
- Am J Clin Nutr-2003-Siekmann-242-9Document8 pagesAm J Clin Nutr-2003-Siekmann-242-9Klinik FiraraNo ratings yet
- Journal Pone 0028812Document8 pagesJournal Pone 0028812Klinik FiraraNo ratings yet
- Int. J. Epidemiol.-2000-Kleinschmidt-355-61Document7 pagesInt. J. Epidemiol.-2000-Kleinschmidt-355-61Klinik FiraraNo ratings yet
- International Journal of Health GeographicsDocument11 pagesInternational Journal of Health GeographicsKlinik FiraraNo ratings yet
- High Affinity Antibodies ToDocument9 pagesHigh Affinity Antibodies ToKlinik FiraraNo ratings yet
- Ferreira 07 ExpParasitolDocument9 pagesFerreira 07 ExpParasitolKlinik FiraraNo ratings yet
- Epidemiology and Spatial Analysis of Malaria in The Northern Peruvian AmazonDocument7 pagesEpidemiology and Spatial Analysis of Malaria in The Northern Peruvian AmazonKlinik FiraraNo ratings yet
- A Spatial Analysis of Individual-And Neighborhood-Level Determinants of Malaria Incidence in Adults, Ontario, CanadaDocument8 pagesA Spatial Analysis of Individual-And Neighborhood-Level Determinants of Malaria Incidence in Adults, Ontario, CanadaKlinik FiraraNo ratings yet
- 1001 FullDocument9 pages1001 FullKlinik FiraraNo ratings yet
- Author Workshop - 2020Document99 pagesAuthor Workshop - 2020rully_mp354No ratings yet
- BCG Scar, Socioeconomic and Nutritional Status A Study ofDocument11 pagesBCG Scar, Socioeconomic and Nutritional Status A Study ofKlinik FiraraNo ratings yet
- Tackling Obesity in ASEAN: Prevalence, Impact, and Guidance On InterventionsDocument79 pagesTackling Obesity in ASEAN: Prevalence, Impact, and Guidance On InterventionsNenden ApriliyantiNo ratings yet
- Nutrients: Obesity: Pathophysiology and InterventionDocument31 pagesNutrients: Obesity: Pathophysiology and Interventionrianza hutasuhutNo ratings yet
- Complications of Obesity in AdultsDocument5 pagesComplications of Obesity in AdultsKlinik FiraraNo ratings yet
- Otolaryngology - Head and Neck Surgery: Practical Guide To Choosing An Appropriate Data DisplayDocument10 pagesOtolaryngology - Head and Neck Surgery: Practical Guide To Choosing An Appropriate Data DisplayKlinik FiraraNo ratings yet
- Orange Grey Medical PresentationDocument5 pagesOrange Grey Medical PresentationKlinik FiraraNo ratings yet
- Public Health Neurological Disorder 2019Document232 pagesPublic Health Neurological Disorder 2019Klinik FiraraNo ratings yet
- Indonesia Healthcare Outlook 2019Document17 pagesIndonesia Healthcare Outlook 2019Klinik Firara100% (1)
- Module V - Neleco2Document7 pagesModule V - Neleco2claudine padillonNo ratings yet
- 4th Quarter P.E. Module 5Document11 pages4th Quarter P.E. Module 5rowena salapiNo ratings yet
- Get in Shape With Caroline Sandry & Health & Fitness MagDocument14 pagesGet in Shape With Caroline Sandry & Health & Fitness MagCaroline Sandry50% (4)
- Tiens Business PlanDocument22 pagesTiens Business PlanSudipta BoseNo ratings yet
- Diabetes NIDocument8 pagesDiabetes NIBeatrizCamposNo ratings yet
- Richa Shah - Thesis AbstractDocument3 pagesRicha Shah - Thesis AbstractRicha ShahNo ratings yet
- Chapter 1 5 Super FinalOther ResearchDocument176 pagesChapter 1 5 Super FinalOther ResearchjeffreyNo ratings yet
- Report Writing Skills ASSIGNMENTDocument4 pagesReport Writing Skills ASSIGNMENTSYED MUHAMMAD USAMA MASOOD50% (2)
- Improving The Physical Health of People With Mental Health ProblemsDocument61 pagesImproving The Physical Health of People With Mental Health ProblemsThierry UhawenimanaNo ratings yet
- AJMC AJP1151 Obesity WebDocument56 pagesAJMC AJP1151 Obesity WebLoc Nguyen TanNo ratings yet
- Eastern Mindoro CollegeDocument4 pagesEastern Mindoro Collegecarmela leonidaNo ratings yet
- Prepare For War by Jamie Lewis PDFDocument62 pagesPrepare For War by Jamie Lewis PDFtom80% (5)
- Public Policy AssignmentDocument6 pagesPublic Policy Assignmentapi-523257212No ratings yet
- Exercise Preferences and ExpectationsDocument277 pagesExercise Preferences and ExpectationsSyed IdrushahabNo ratings yet
- A Framework For Mission-Oriented InnovationDocument16 pagesA Framework For Mission-Oriented InnovationaryNo ratings yet
- Research Paper On Family Mealtime WeeblyDocument7 pagesResearch Paper On Family Mealtime Weeblyapi-239973060No ratings yet
- Smart Fridge Artle HCIDocument12 pagesSmart Fridge Artle HCIvipgiggles2010No ratings yet
- Science Folio FINALESTDocument13 pagesScience Folio FINALESTDesNo ratings yet
- M.Sc. Semester IV Syllabus DISTRIBUTIONDocument16 pagesM.Sc. Semester IV Syllabus DISTRIBUTIONSuvNo ratings yet
- National pediatric evaluationDocument8 pagesNational pediatric evaluationMaya Susanti100% (1)
- Test Bank For Nutrition Diet Therapy 10th Edition Ruth A RothDocument10 pagesTest Bank For Nutrition Diet Therapy 10th Edition Ruth A RothAmber Gault100% (24)
- Writting EassayDocument58 pagesWritting EassayLinkon PeterNo ratings yet
- Grade 10 MapehDocument2 pagesGrade 10 MapehMae Ann Ebitner EspañolNo ratings yet
- The Lyon Protocol Dr. Gabrielle LyonDocument20 pagesThe Lyon Protocol Dr. Gabrielle LyonIvan Farkaš100% (2)
- Waist CircumferenceDocument7 pagesWaist CircumferenceYohanes Denny TanugerahNo ratings yet
- 21 Life Lessons From Livin La Vida Low CarbDocument370 pages21 Life Lessons From Livin La Vida Low CarbKathryn EvansNo ratings yet
- Plenary Session. 1. Challenges-Opportunity of CVD Services in Universal Coverage. Dr. Anwar Santoso SPJPKDocument32 pagesPlenary Session. 1. Challenges-Opportunity of CVD Services in Universal Coverage. Dr. Anwar Santoso SPJPKannisNo ratings yet
- Arginine Metabolism and its Role in Growth, Health and DiseaseDocument26 pagesArginine Metabolism and its Role in Growth, Health and Diseaseאנריקה שטרמר דיאטן קליניNo ratings yet
- Fast Food Consumption Trend and its Effect on Pakistani SocietyDocument9 pagesFast Food Consumption Trend and its Effect on Pakistani SocietyAhmed SaeedNo ratings yet
- 7e's Lesson Plan For Physical EducationDocument3 pages7e's Lesson Plan For Physical EducationMajelyn A. Icalina100% (3)