You might also like
- Interrelationship Between Carbohydrate Protein Fat MetabolismDocument12 pagesInterrelationship Between Carbohydrate Protein Fat MetabolismShailaja Neaupane100% (5)
- Glucose regulation-RSKDocument4 pagesGlucose regulation-RSKdevilalshingh9525No ratings yet
- Endocrine Pancreas & Fuel Homeostasis: Learning ObjectivesDocument7 pagesEndocrine Pancreas & Fuel Homeostasis: Learning ObjectivesMaggieLockeNo ratings yet
- CHO Lec 1Document9 pagesCHO Lec 1sajjadany50No ratings yet
- Basics in Clinical Nutrition: Carbohydrate Metabolism: Luc TappyDocument4 pagesBasics in Clinical Nutrition: Carbohydrate Metabolism: Luc Tappymarliana100% (1)
- Fisiopatologia de La Crisis HipoglicemicaDocument11 pagesFisiopatologia de La Crisis HipoglicemicaLemur DaJuliNo ratings yet
- Signaling Pathways of Glucose in Our Body - PART ONEDocument9 pagesSignaling Pathways of Glucose in Our Body - PART ONEMaria Jose MartinezNo ratings yet
- Post Lab Question Experiment 5Document6 pagesPost Lab Question Experiment 5Skefadiuto100% (3)
- Pancreatic Hormones Regulate Blood GlucoseDocument10 pagesPancreatic Hormones Regulate Blood Glucosekrizzia raymundoNo ratings yet
- Endocrine Pancreas & Fuel Homeostasis: Learning ObjectivesDocument7 pagesEndocrine Pancreas & Fuel Homeostasis: Learning ObjectivesEmmanuel NhandaraNo ratings yet
- 1808Document14 pages180811. Nguyễn Võ Như HuỳnhNo ratings yet
- PresentationDocument37 pagesPresentationFaizan Asif045No ratings yet
- Gluco Neo GenesisDocument6 pagesGluco Neo GenesisNizam YahyaNo ratings yet
- Notes On CHO MetabDocument6 pagesNotes On CHO Metaboddone_out100% (1)
- Chapter 5 Glucose HomeostasisDocument15 pagesChapter 5 Glucose Homeostasistliviu334066No ratings yet
- Book Chapter Carbohydrate Storage and MetabolismDocument16 pagesBook Chapter Carbohydrate Storage and Metabolismq8laNo ratings yet
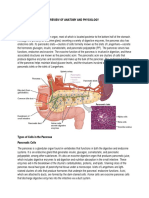
- Review of Anatomy and PhysiologyDocument7 pagesReview of Anatomy and PhysiologyKyla CalzadoNo ratings yet
- Anatomy and PhysiologyDocument2 pagesAnatomy and PhysiologyJeffrey Calicdan BucalaNo ratings yet
- Metabolism Dysorder and Carbohydrate DigestionDocument44 pagesMetabolism Dysorder and Carbohydrate DigestionPrajna PNo ratings yet
- Blood Sugar Reg-WPS OfficeDocument8 pagesBlood Sugar Reg-WPS Officetanjidul islamNo ratings yet
- pansreas.met.mod._aa12f2dfff68e85ba25058e03a20fb9aDocument19 pagespansreas.met.mod._aa12f2dfff68e85ba25058e03a20fb9aLT DRAGONXNo ratings yet
- Physiologic Response To Hypoglycemia in Healthy Individuals and Patients With Diabetes MellitusDocument14 pagesPhysiologic Response To Hypoglycemia in Healthy Individuals and Patients With Diabetes Mellitusmayteveronica1000No ratings yet
- Homeostatis Principles and Regulation of Glucose in BloodDocument13 pagesHomeostatis Principles and Regulation of Glucose in Bloodk75544863No ratings yet
- Irregularity of Glycogen Synthase Homeostasis Complications Hepatic and Neuro DefectsDocument11 pagesIrregularity of Glycogen Synthase Homeostasis Complications Hepatic and Neuro DefectsEditor IJTSRDNo ratings yet
- Gluconeogenesis & Glycogen 2011Document35 pagesGluconeogenesis & Glycogen 2011glenn johnstonNo ratings yet
- Presentation NBCDocument6 pagesPresentation NBCD. Akansha GoudNo ratings yet
- Endo Pancrease ElhDocument27 pagesEndo Pancrease Elhodiodi57No ratings yet
- Costa Rica GlucoseDocument91 pagesCosta Rica GlucoseAnthony HarderNo ratings yet
- Anaphy PancreasDocument6 pagesAnaphy PancreasAELYN SEVILLANo ratings yet
- P4-Obesity and The Regulation of Body MassDocument8 pagesP4-Obesity and The Regulation of Body MassOcta RenitaNo ratings yet
- Glucose John SantangeloDocument45 pagesGlucose John Santangelosaint5470No ratings yet
- InsulinDocument50 pagesInsulinTandin SonamNo ratings yet
- Blood Glucose Regulation and HormonesDocument9 pagesBlood Glucose Regulation and HormonesJohn OgunsolaNo ratings yet
- 4-CHO Metabolism 2 & DMDocument33 pages4-CHO Metabolism 2 & DMlobna adelNo ratings yet
- 5.0 Hormonal Control of Carbohydrate MetabolismDocument10 pages5.0 Hormonal Control of Carbohydrate Metabolismeka saputraNo ratings yet
- Clinical Chemistry - CarbohydratesDocument8 pagesClinical Chemistry - Carbohydratesrosellae.No ratings yet
- Insulin, Glucagon, and Diabetes Mellitus ExplainedDocument22 pagesInsulin, Glucagon, and Diabetes Mellitus Explainedghadeer1No ratings yet
- Major pathways of carbohydrate metabolismDocument119 pagesMajor pathways of carbohydrate metabolismsimasNo ratings yet
- Insulin, Glucagon, and Diabetes MellitusDocument22 pagesInsulin, Glucagon, and Diabetes Mellitusghadeer1No ratings yet
- 05.17 Metabolic Effects of Insulin and Glucagon - W2016-2Document28 pages05.17 Metabolic Effects of Insulin and Glucagon - W2016-2Rima Gi100% (1)
- Bab Ii PDFDocument14 pagesBab Ii PDFyevanharrybrataNo ratings yet
- محاضرات الكيمياء كاملة PDFDocument124 pagesمحاضرات الكيمياء كاملة PDFفراس الموسويNo ratings yet
- Wallah THAK GI ME?Document5 pagesWallah THAK GI ME?Usman EngNo ratings yet
- Physio 7Document12 pagesPhysio 7amjadNo ratings yet
- Regulation of MetabolismDocument42 pagesRegulation of Metabolismامجد حسين جواد كاظمNo ratings yet
- Metabolism - Glycolysis-2Document8 pagesMetabolism - Glycolysis-2Wendy SierraNo ratings yet
- Glucagon ReleaseDocument10 pagesGlucagon Releasesilvia pradoNo ratings yet
- Functions of CarbohydrateDocument11 pagesFunctions of CarbohydrateAbdulbaasit AyoolaNo ratings yet
- Kortizol FizyolojiDocument5 pagesKortizol FizyolojiVedat KacarNo ratings yet
- Pancreas: Cells and Secretions of The Pancreatic IsletsDocument8 pagesPancreas: Cells and Secretions of The Pancreatic IsletsansahNo ratings yet
- Diabetic Emergencies in Small AnimalsDocument17 pagesDiabetic Emergencies in Small Animalsdia_dianneNo ratings yet
- Pancreatic Islets Hormones Regulate Blood GlucoseDocument4 pagesPancreatic Islets Hormones Regulate Blood GlucoseSophia OcayNo ratings yet
- Blood Sugar: AtionDocument93 pagesBlood Sugar: Ationmidhunramesh007100% (2)
- 11. Hypoglycemia and hyperglycemia GM eng 2022Document3 pages11. Hypoglycemia and hyperglycemia GM eng 2022kabulkabulovich5No ratings yet
- Diabetes Spectr 2004 Aronoff 183 90Document8 pagesDiabetes Spectr 2004 Aronoff 183 90Bodea DoruNo ratings yet
- Intergration of MetabolismDocument112 pagesIntergration of MetabolismSantino MajokNo ratings yet
- Lec 11 - Insulin and Glucagon PDFDocument17 pagesLec 11 - Insulin and Glucagon PDFrajeshNo ratings yet
- Insulin and Glucagon HormonesDocument42 pagesInsulin and Glucagon HormonesRoseNo ratings yet
- Insulin and Glucagon Regulation of Metabolic HomeostasisDocument40 pagesInsulin and Glucagon Regulation of Metabolic HomeostasisEka HabinaNo ratings yet
- Hypoglycemia, A Simple Guide To The Condition, Treatment And Related ConditionsFrom EverandHypoglycemia, A Simple Guide To The Condition, Treatment And Related ConditionsNo ratings yet
- Screenshot 2021-06-23 at 11.36.48 AMDocument116 pagesScreenshot 2021-06-23 at 11.36.48 AMSmitha ShekarNo ratings yet
- Circulatory SystemDocument14 pagesCirculatory Systemsmbdy tbhhhNo ratings yet
- Female Reproductive Anatomy GuideDocument6 pagesFemale Reproductive Anatomy GuideanojanNo ratings yet
- EMT Refresher Practice Exam ReviewDocument25 pagesEMT Refresher Practice Exam ReviewNimra NazeerNo ratings yet
- Jurnal Biosel Serial Endosymbiosis TheoryDocument4 pagesJurnal Biosel Serial Endosymbiosis TheoryHeny SinagaNo ratings yet
- Histology of SkinDocument33 pagesHistology of SkinH R A K Kulathilaka75% (4)
- Test Maternal and PediaDocument11 pagesTest Maternal and PediaDefensor Pison GringgoNo ratings yet
- Section: General ObstetricsDocument15 pagesSection: General ObstetricsvarmaNo ratings yet
- Mechanical Vectors of Medical ImportanceDocument35 pagesMechanical Vectors of Medical ImportanceCLEMENTNo ratings yet
- Endocrine Physiology LaboratoryDocument1 pageEndocrine Physiology LaboratoryRizza Mae Salvania100% (1)
- 10-2019 - Canine Tactical Combat Casualty Care (CTCCC)Document17 pages10-2019 - Canine Tactical Combat Casualty Care (CTCCC)pibulinNo ratings yet
- CNS ExaminationDocument3 pagesCNS ExaminationsekharNo ratings yet
- Danielle Karen Widjaja 22010113130175 Lap - Kti Bab2Document2 pagesDanielle Karen Widjaja 22010113130175 Lap - Kti Bab2Mamanya Fadhil HaniNo ratings yet
- Fetuin-A Levels in Hyperthyroidism.Document5 pagesFetuin-A Levels in Hyperthyroidism.yekimaNo ratings yet
- Final Defense (RVSD)Document37 pagesFinal Defense (RVSD)Torres Ken Robin DeldaNo ratings yet
- Callus CultureDocument24 pagesCallus CultureHabiba Majeed MalikNo ratings yet
- My Journey With Mum & MDSDocument54 pagesMy Journey With Mum & MDSIlene OngNo ratings yet
- Living Organisms CharacteristicsDocument6 pagesLiving Organisms CharacteristicslewisNo ratings yet
- Human Health and DiseasesDocument24 pagesHuman Health and DiseasesAnujith AnuNo ratings yet
- Test Bank For The Developing Human Clinically Oriented Embryology 8th Edition Moore Isbn 10 1416037063 Isbn 13 9781416037064Document11 pagesTest Bank For The Developing Human Clinically Oriented Embryology 8th Edition Moore Isbn 10 1416037063 Isbn 13 9781416037064Helen Richardson100% (29)
- Interpreting The Chest Radio Graph Friendly)Document9 pagesInterpreting The Chest Radio Graph Friendly)Vagner BorgesNo ratings yet
- Es Racp Dwe Adult Medical Sciences 101 170Document26 pagesEs Racp Dwe Adult Medical Sciences 101 170Gurki BajwaNo ratings yet
- By Saifullah Khalid (Lecturer) School of Physiotherapy, IPM&R, Dow University of Health Sciences, KarachiDocument31 pagesBy Saifullah Khalid (Lecturer) School of Physiotherapy, IPM&R, Dow University of Health Sciences, KarachiAbhinandan RamkrishnanNo ratings yet
- Nursing Care Plan For Postpartum HemorrhageDocument2 pagesNursing Care Plan For Postpartum HemorrhageDianne Mae100% (1)
- New Estrogen and ProgesteroneDocument39 pagesNew Estrogen and ProgesteroneWegrimel AriegaraNo ratings yet
- All About SharksDocument3 pagesAll About SharksGutmonarchNo ratings yet
- Muscle Relaxants Classification and Mechanisms of ActionDocument22 pagesMuscle Relaxants Classification and Mechanisms of Actionvinay0717100% (1)
- Biol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairDocument6 pagesBiol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairJohn Paradise100% (1)
- Approach To Peripheral NeuropathyDocument56 pagesApproach To Peripheral NeuropathyRhomizal MazaliNo ratings yet