You might also like
- NIH Public Access: Author ManuscriptDocument28 pagesNIH Public Access: Author ManuscriptAlfonso LemaNo ratings yet
- Circulating Myeloid-Derived MMP8 in Stress Susceptibility and DepressionDocument35 pagesCirculating Myeloid-Derived MMP8 in Stress Susceptibility and Depressionnishantbhardwaj104No ratings yet
- jcm-08-00537 YDA 7Document11 pagesjcm-08-00537 YDA 7Muhammad Sandy Ali YafieNo ratings yet
- Srep 45623Document14 pagesSrep 45623javi222222No ratings yet
- Nhsjs SubmissionDocument8 pagesNhsjs Submissionapi-410186016No ratings yet
- Artigo 2 Baixado de NovoDocument7 pagesArtigo 2 Baixado de NovofefaegusNo ratings yet
- Cortical Mechanisms in MigraineDocument14 pagesCortical Mechanisms in MigraineLoida CamargoNo ratings yet
- RetrieveDocument12 pagesRetrievei.ch.lampropoulosNo ratings yet
- 2017 Resilience and Vulnerability To Pain and Inflammation in The HippocampusDocument14 pages2017 Resilience and Vulnerability To Pain and Inflammation in The HippocampusCristian OyarzoNo ratings yet
- Reich DS - MSDocument17 pagesReich DS - MSinesNo ratings yet
- Neuroinflammation: The Devil Is in The DetailsDocument18 pagesNeuroinflammation: The Devil Is in The DetailsAkriti NoNo ratings yet
- Immune System Connection To The CNSDocument12 pagesImmune System Connection To The CNSAndrew GrantNo ratings yet
- Mesenchymal Stem Cells For Spinal Cord Injury: Current Options, Limitations, and Future of Cell TherapyDocument26 pagesMesenchymal Stem Cells For Spinal Cord Injury: Current Options, Limitations, and Future of Cell Therapyadlin amaliaNo ratings yet
- Ladecola C, Anrather J. (2012)Document13 pagesLadecola C, Anrather J. (2012)Rusda AnandaNo ratings yet
- Cellular Biology of Fracture HealingDocument16 pagesCellular Biology of Fracture HealingNadira NurinNo ratings yet
- Redox BiologyDocument14 pagesRedox BiologyLin LINo ratings yet
- Mechanisms Underlying Brain Aging Under Normal and Pathological ConditionsDocument12 pagesMechanisms Underlying Brain Aging Under Normal and Pathological ConditionsDiana BernalNo ratings yet
- Fisiop Lesion Traumatic ADocument16 pagesFisiop Lesion Traumatic ADaniel SanchezNo ratings yet
- Inflammatory Response After Neurosurgery PDFDocument18 pagesInflammatory Response After Neurosurgery PDFnetim69No ratings yet
- 1 s2.0 S0896627314001718 MainDocument18 pages1 s2.0 S0896627314001718 MainAhmet KayalıNo ratings yet
- Macrophage DualityDocument13 pagesMacrophage DualityzotzedNo ratings yet
- Liu 等。 - 2021 - Extracellular Vesicles Derived From M2 Microglia rDocument14 pagesLiu 等。 - 2021 - Extracellular Vesicles Derived From M2 Microglia rYouPingLiNo ratings yet
- Andrade 2016Document12 pagesAndrade 2016myztNo ratings yet
- A 45-Year Old Male With Left-Sided Hemihypesthesia: Clinical HistoryDocument4 pagesA 45-Year Old Male With Left-Sided Hemihypesthesia: Clinical HistorywillygopeNo ratings yet
- Mekanisme PDFDocument9 pagesMekanisme PDFAndyk Strapilococus AureusNo ratings yet
- Nature 14432Document17 pagesNature 14432LUISNo ratings yet
- A 10-Year Old Girl With Neck Pain: Clinical HistoryDocument4 pagesA 10-Year Old Girl With Neck Pain: Clinical HistorywillygopeNo ratings yet
- Post-Ischemic in Ammation in The Brain: Takashi Shichita, Ryota Sakaguchi, Mayu Suzuki and Akihiko YoshimuraDocument7 pagesPost-Ischemic in Ammation in The Brain: Takashi Shichita, Ryota Sakaguchi, Mayu Suzuki and Akihiko YoshimuraAlex GasnasNo ratings yet
- A Neuron's PerspectiveDocument15 pagesA Neuron's PerspectivePiali ChatterjeeNo ratings yet
- Histopathological Changes in Acute Ischemic Stroke: R I G I N A L A P E RDocument13 pagesHistopathological Changes in Acute Ischemic Stroke: R I G I N A L A P E RJosé Luis García MezaNo ratings yet
- Savitz Et Al-2016-Stem CellsDocument6 pagesSavitz Et Al-2016-Stem CellsMaha Satya Dwi PalgunaNo ratings yet
- Lee 2008Document14 pagesLee 2008Laura Duarte RojasNo ratings yet
- Biologia Celular en Consolidación ÓseaDocument16 pagesBiologia Celular en Consolidación Óseadaniela rojasNo ratings yet
- NeurologyDocument19 pagesNeurologymerosaber622001No ratings yet
- Local Immunomodulation and Muscle Progenitor Cells Induce Recovery in Atrophied Muscles in Spinal Cord Injury PatientsDocument10 pagesLocal Immunomodulation and Muscle Progenitor Cells Induce Recovery in Atrophied Muscles in Spinal Cord Injury PatientsGustavo MovigliaNo ratings yet
- Development of Monocytes, Macrophages and Dendritic CellsDocument12 pagesDevelopment of Monocytes, Macrophages and Dendritic CellsHelenaNo ratings yet
- Badaut Zhang Preface2019 ADocument5 pagesBadaut Zhang Preface2019 AGonzalo AltamiranoNo ratings yet
- Behav - Brain - Res - 2012 - DRKFDocument7 pagesBehav - Brain - Res - 2012 - DRKFFarah AlshammasiNo ratings yet
- Asahi 2001Document9 pagesAsahi 2001MariaNo ratings yet
- DC 1 PDFDocument25 pagesDC 1 PDFRebecca SandoicaNo ratings yet
- Antifungal Drug Miconazole Ameliorated Memory Deficits in A Mouse Model of LPS-Induced Memory Loss Through Targeting INOSDocument14 pagesAntifungal Drug Miconazole Ameliorated Memory Deficits in A Mouse Model of LPS-Induced Memory Loss Through Targeting INOSSemper PhiNo ratings yet
- 2604 6180 RMN 21 3 111Document11 pages2604 6180 RMN 21 3 111NICOL DAYANNA GONZALEZ OSMANo ratings yet
- Neurogenic Arthrogryposis and The Power of Phenoty 2021 Neuromuscular DisordDocument8 pagesNeurogenic Arthrogryposis and The Power of Phenoty 2021 Neuromuscular DisordSuzie Simone Mardones SilvaNo ratings yet
- Neurogenic Arthrogryposis and The Power of Phenoty - 2021 - Neuromuscular DisordDocument8 pagesNeurogenic Arthrogryposis and The Power of Phenoty - 2021 - Neuromuscular DisordSuzie Simone Mardones SilvaNo ratings yet
- Neuro-Endocrine Networks Controlling Immune System in Health and DiseaseDocument10 pagesNeuro-Endocrine Networks Controlling Immune System in Health and DiseaseUmmu AthiyyahNo ratings yet
- Treatment and Evaluation Advances in Leprosy Neuropathy: Gigi J. Ebenezer David M. ScollardDocument14 pagesTreatment and Evaluation Advances in Leprosy Neuropathy: Gigi J. Ebenezer David M. ScollardSisca NataliaNo ratings yet
- Mesenteric Stem Cell For Central Nervous System LesionDocument15 pagesMesenteric Stem Cell For Central Nervous System LesionGlen Sandy SaapangNo ratings yet
- 5437 FullDocument14 pages5437 FullFahrunnisa NurdinNo ratings yet
- Neuroscience: Archives ofDocument6 pagesNeuroscience: Archives ofCom DigfulNo ratings yet
- Neuroscience: Archives ofDocument6 pagesNeuroscience: Archives ofCom DigfulNo ratings yet
- Brain Immunology and Immunotherapy in Brain TumoursDocument14 pagesBrain Immunology and Immunotherapy in Brain TumoursAnonymous QLadTClydkNo ratings yet
- 5 Is - Central - Nervous - System - An - Immune-Privileged - SitDocument7 pages5 Is - Central - Nervous - System - An - Immune-Privileged - SitAna Maria Vargas RuizNo ratings yet
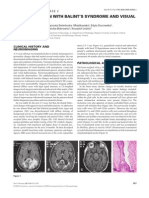
- 31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingDocument4 pages31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingwillygopeNo ratings yet
- Interactions Between Epilepsy and Plasticity: PharmaceuticalsDocument18 pagesInteractions Between Epilepsy and Plasticity: PharmaceuticalsYahya Idriss IbrhimNo ratings yet
- The Shrinking Brain: Cerebral Atrophy Following Traumatic Brain InjuryDocument19 pagesThe Shrinking Brain: Cerebral Atrophy Following Traumatic Brain InjuryidkNo ratings yet
- Regulation of Axonal Regeneration After Mammalian Spinal Cord InjuryDocument18 pagesRegulation of Axonal Regeneration After Mammalian Spinal Cord Injuryelif.onsozNo ratings yet
- NeuroiflammationDocument9 pagesNeuroiflammationGabrielaNo ratings yet
- Neutrophils at The Crossroads of Innate and Adaptive Immunity. J Leukoc Biol. 2020 JulDocument20 pagesNeutrophils at The Crossroads of Innate and Adaptive Immunity. J Leukoc Biol. 2020 Julantonychino10No ratings yet
- Telaah 3Document25 pagesTelaah 3anri albarruNo ratings yet
- Detection and Quantification of Bovine Papillomavirus DNA by Digital Droplet PCR in Sheep BloodDocument7 pagesDetection and Quantification of Bovine Papillomavirus DNA by Digital Droplet PCR in Sheep Bloodmuralim_phy1986No ratings yet
- A Material Stress Test Study On Occurrence of Leakage and Material Failure of Peritoneal Dialysis (PD) CathetersDocument6 pagesA Material Stress Test Study On Occurrence of Leakage and Material Failure of Peritoneal Dialysis (PD) Cathetersmuralim_phy1986No ratings yet
- Wild Bornean Orangutans Experience Muscle Catabolism During Episodes of Fruit ScarcityDocument7 pagesWild Bornean Orangutans Experience Muscle Catabolism During Episodes of Fruit Scarcitymuralim_phy1986No ratings yet
- NFL Strongly Correlates With TNF R1 in The Plasma of AD Patients, But Not With Cognitive DeclineDocument7 pagesNFL Strongly Correlates With TNF R1 in The Plasma of AD Patients, But Not With Cognitive Declinemuralim_phy1986No ratings yet
- Histidine Rich Glycoprotein As A Prognostic Biomarker For SepsisDocument7 pagesHistidine Rich Glycoprotein As A Prognostic Biomarker For Sepsismuralim_phy1986No ratings yet
- Remarkable Plasma Resistance Performance by Nanocrystalline Y O Mgo Composite Ceramics For Semiconductor Industry ApplicationsDocument7 pagesRemarkable Plasma Resistance Performance by Nanocrystalline Y O Mgo Composite Ceramics For Semiconductor Industry Applicationsmuralim_phy1986No ratings yet
- SIGLEC1 (CD169) : A Marker of Active Neuroinflammation in The Brain But Not in The Blood of Multiple Sclerosis PatientsDocument8 pagesSIGLEC1 (CD169) : A Marker of Active Neuroinflammation in The Brain But Not in The Blood of Multiple Sclerosis Patientsmuralim_phy1986No ratings yet
- Process Optimization For The Supercritical Carbondioxide Extraction of Lycopene From Ripe Grapefruit (Citrus Paradisi) EndocarpDocument8 pagesProcess Optimization For The Supercritical Carbondioxide Extraction of Lycopene From Ripe Grapefruit (Citrus Paradisi) Endocarpmuralim_phy1986No ratings yet
- Zonally Asymmetric Phytoplankton Response To The Southern Annular Mode in The Marginal Sea of The Southern OceanDocument8 pagesZonally Asymmetric Phytoplankton Response To The Southern Annular Mode in The Marginal Sea of The Southern Oceanmuralim_phy1986No ratings yet
- SrAlNbO6 EtcDocument10 pagesSrAlNbO6 Etcmuralim_phy1986No ratings yet
- Improved Efficiency of Coding Systems With Health Information TechnologyDocument6 pagesImproved Efficiency of Coding Systems With Health Information Technologymuralim_phy1986No ratings yet
- Comsol-Based Modeling and Simulation of Sno /rgo Gas Sensor For Detection of NoDocument12 pagesComsol-Based Modeling and Simulation of Sno /rgo Gas Sensor For Detection of Nomuralim_phy1986No ratings yet
- Long Term Results of Sirolimus Treatment in Lymphangioleiomyomatosis: A Single Referral Centre ExperienceDocument9 pagesLong Term Results of Sirolimus Treatment in Lymphangioleiomyomatosis: A Single Referral Centre Experiencemuralim_phy1986No ratings yet
- Quantum Entanglement and Statistics of Photons On A Beam Splitter in The Form of Coupled WaveguidesDocument7 pagesQuantum Entanglement and Statistics of Photons On A Beam Splitter in The Form of Coupled Waveguidesmuralim_phy1986No ratings yet
- Inhibition of Checkpoint Kinase 1 Potentiates Anticancer Activity of Gemcitabine in Bladder Cancer CellsDocument8 pagesInhibition of Checkpoint Kinase 1 Potentiates Anticancer Activity of Gemcitabine in Bladder Cancer Cellsmuralim_phy1986No ratings yet
- Gender Differences in Health Related Quality of Life Measured by The Sarcoidosis Health QuestionnaireDocument7 pagesGender Differences in Health Related Quality of Life Measured by The Sarcoidosis Health Questionnairemuralim_phy1986No ratings yet
- Elevated Transaminases and Hypoalbuminemia in Covid 19 Are Prognostic Factors For Disease SeverityDocument5 pagesElevated Transaminases and Hypoalbuminemia in Covid 19 Are Prognostic Factors For Disease Severitymuralim_phy1986No ratings yet
- Daily Variation of Macronutrient Concentrations in Mature Human Milk Over 3 WeeksDocument9 pagesDaily Variation of Macronutrient Concentrations in Mature Human Milk Over 3 Weeksmuralim_phy1986No ratings yet
- Surge of Severe Acute Respiratory Syndrome Coronavirus 2 Infections Linked To Single Introduction of A Virus Strain in Myanmar, 2020Document6 pagesSurge of Severe Acute Respiratory Syndrome Coronavirus 2 Infections Linked To Single Introduction of A Virus Strain in Myanmar, 2020muralim_phy1986No ratings yet
- Serum Alanine Aminotransferase As An Early Marker of Outcomes in Patients Receiving Anti PD 1 or Anti CTLA 4 AntibodyDocument10 pagesSerum Alanine Aminotransferase As An Early Marker of Outcomes in Patients Receiving Anti PD 1 or Anti CTLA 4 Antibodymuralim_phy1986No ratings yet
- s41598 021 89712 4Document7 pagess41598 021 89712 4muralim_phy1986No ratings yet
- Divergent Nucleic Acid Allocation in Juvenile Insects of Different Metamorphosis ModesDocument8 pagesDivergent Nucleic Acid Allocation in Juvenile Insects of Different Metamorphosis Modesmuralim_phy1986No ratings yet
- Comsol-Based Modeling and Simulation of Sno /rgo Gas Sensor For Detection of NoDocument12 pagesComsol-Based Modeling and Simulation of Sno /rgo Gas Sensor For Detection of Nomuralim_phy1986No ratings yet
- Quantitative Determination of Spin-Orbit Induced Magnetic Field in Gamnas by Field Scan Planar Hall MeasurementsDocument8 pagesQuantitative Determination of Spin-Orbit Induced Magnetic Field in Gamnas by Field Scan Planar Hall Measurementsmuralim_phy1986No ratings yet
- GrandmasHouseP2 0.37 - Walkthrough Rev 1.15Document24 pagesGrandmasHouseP2 0.37 - Walkthrough Rev 1.15kunwarpiya040893No ratings yet
- Guideline Perkeni 2019Document29 pagesGuideline Perkeni 2019Tiens MonisaNo ratings yet
- Caregiving 12 Module 1 4th QuarterDocument23 pagesCaregiving 12 Module 1 4th QuarterBethuel AlquirozNo ratings yet
- MESS RISK ASSESMENT REPORT - Group 1 - D PDFDocument12 pagesMESS RISK ASSESMENT REPORT - Group 1 - D PDFSurajNo ratings yet
- Time To Synergize Mental Health With Brain Health: CommentDocument3 pagesTime To Synergize Mental Health With Brain Health: Commentsoliveira_149796No ratings yet
- APLEY & SOLOMON'S - 5. OsteoarthritisDocument19 pagesAPLEY & SOLOMON'S - 5. OsteoarthritisOngko SetunggalNo ratings yet
- Public Health ConceptsDocument9 pagesPublic Health ConceptsSucheta MitraNo ratings yet
- Dr. Shanthi Rengarajan V The Oriental Insurance CompanyDocument12 pagesDr. Shanthi Rengarajan V The Oriental Insurance Companyparvati anilkumarNo ratings yet
- v1 - Planets of Orthodontics - Vi - Essentials of Orthodontics 1Document93 pagesv1 - Planets of Orthodontics - Vi - Essentials of Orthodontics 1PhanQuangHuyNo ratings yet
- Index No Pvms No Item NameDocument50 pagesIndex No Pvms No Item NamePcrNo ratings yet
- امتحان شامل -أداة قياس المهارات العملية - 1Document1 pageامتحان شامل -أداة قياس المهارات العملية - 1Rn nadeenNo ratings yet
- Hope - 1 Grade 11: Exercise For FitnessDocument6 pagesHope - 1 Grade 11: Exercise For FitnessTABBAL JAN PAOLONo ratings yet
- Bing Kls 8Document5 pagesBing Kls 8Soni DwinugrohoNo ratings yet
- 10 HodobaDocument14 pages10 HodobaHepicentar NišaNo ratings yet
- Updated BMI Monitoring of Pat Dexter L Ginon 1Document3 pagesUpdated BMI Monitoring of Pat Dexter L Ginon 1Fredimar PatarayNo ratings yet
- Ibong Adarna - Summary English VersionDocument7 pagesIbong Adarna - Summary English Versiontin.calli26No ratings yet
- Genre Analysis Outlining Worksheet 1 1Document4 pagesGenre Analysis Outlining Worksheet 1 1api-733516026No ratings yet
- Kyoto OjoDocument2 pagesKyoto OjoFernando SanNo ratings yet
- Phase IDocument28 pagesPhase INandagopal KNo ratings yet
- Does This Dyspneic Patient in The Emergency Department Have Congestive Heart Failure?Document13 pagesDoes This Dyspneic Patient in The Emergency Department Have Congestive Heart Failure?Jorge MéndezNo ratings yet
- Portfolio of Products For ExportDocument8 pagesPortfolio of Products For ExportYeni Paola Ibarra RodríguezNo ratings yet
- Compensation & Reward ManagementDocument227 pagesCompensation & Reward ManagementjitendersharmajiNo ratings yet
- Uterine Fibroid-IzzatDocument20 pagesUterine Fibroid-IzzatcopperNo ratings yet
- Lesson-1 Understanding Consumer MotivationDocument2 pagesLesson-1 Understanding Consumer MotivationKristine Angela RanotNo ratings yet
- Preassessment: Setting Up The Classroom For Epp/TleDocument21 pagesPreassessment: Setting Up The Classroom For Epp/TleMhazzy ReyesNo ratings yet
- Tpl-Pcipl JV: Operational Control Procedure For Hard Rock BlastingDocument7 pagesTpl-Pcipl JV: Operational Control Procedure For Hard Rock BlastingNirbhay KumarNo ratings yet
- Jihan Rahma Fadillah 46-50Document14 pagesJihan Rahma Fadillah 46-50Hendry RahmanantoNo ratings yet
- Smart Medical Card SynopsisDocument6 pagesSmart Medical Card SynopsisYashika ChouhanNo ratings yet
- Health Insurance Guidelines 2020-21Document25 pagesHealth Insurance Guidelines 2020-21NIRAJ BAHEKARNo ratings yet
- Urinary Retention NCP GARCIA J.Document2 pagesUrinary Retention NCP GARCIA J.Jimlord GarciaNo ratings yet