You might also like
- The Four Common Types of Parenting StylesDocument11 pagesThe Four Common Types of Parenting StylesIka_Dyah_Purwa_1972100% (3)
- Wrap Book A4Document27 pagesWrap Book A4doscribe100% (1)
- Mcqs of Family LawDocument24 pagesMcqs of Family LawVenugopal Mantraratnam32% (19)
- Aircraft Instruments and Avionics PDFDocument211 pagesAircraft Instruments and Avionics PDFairbuk doeing88% (8)
- The Strange ICA Stones of PeruDocument31 pagesThe Strange ICA Stones of PeruRoman Aleshkevich100% (1)
- Form Ibpr Haul Road PDFDocument2 pagesForm Ibpr Haul Road PDFelvandi100% (1)
- Supra 644, 744, 844 Beginning With Serial# Hfy90593608 - Sunbelt ...Document68 pagesSupra 644, 744, 844 Beginning With Serial# Hfy90593608 - Sunbelt ...Glacial100% (1)
- BD 1050 PartesDocument26 pagesBD 1050 PartesFelipe RojasNo ratings yet
- Effect of Substrate and Rearing Period On The Yield and Quality of Housefly Larvae ( (Musca Domestica) As Protein Ingredient in Fish DietsDocument4 pagesEffect of Substrate and Rearing Period On The Yield and Quality of Housefly Larvae ( (Musca Domestica) As Protein Ingredient in Fish DietsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Genotyping and Phenotyping Strategies For Genetic Improvement of Meat Quality and Carcass Composition in SwineDocument16 pagesGenotyping and Phenotyping Strategies For Genetic Improvement of Meat Quality and Carcass Composition in Swinestephano portugalNo ratings yet
- Genomic Prediction Low Density Markers AquacultureDocument8 pagesGenomic Prediction Low Density Markers AquaculturetrongtqNo ratings yet
- Genome Editing Livestock Breeding ProgramsDocument17 pagesGenome Editing Livestock Breeding Programsmirielys.rondaNo ratings yet
- 2022-OCS_BSDocument10 pages2022-OCS_BSDubravko ŠkorputNo ratings yet
- Abdelqader Et - Al., 2007, Characterization of Local Chicken Under Different Production and Management of JordanDocument11 pagesAbdelqader Et - Al., 2007, Characterization of Local Chicken Under Different Production and Management of JordanAlok BhartiNo ratings yet
- Estimation of Genetic and Phenotypic Parameters For Sow Productivity Traits in South African Large White PigsDocument9 pagesEstimation of Genetic and Phenotypic Parameters For Sow Productivity Traits in South African Large White Pigsonurb90No ratings yet
- Comparison of Differente Breeding Strategies To Improve Alpaca Fibre Prodution in A Low-Input Production SystemDocument13 pagesComparison of Differente Breeding Strategies To Improve Alpaca Fibre Prodution in A Low-Input Production SystemEdu Barragan HanccoNo ratings yet
- Multi-breed genomic predictions and functional variants for fertility of tropical bullsDocument19 pagesMulti-breed genomic predictions and functional variants for fertility of tropical bullsBahlibiNo ratings yet
- Statistical Methods For Plant BreedingDocument31 pagesStatistical Methods For Plant BreedingIrena CalicNo ratings yet
- Livestock Science: P. Herold, R. Roessler, A. Willam, H. Momm, A. Valle ZárateDocument10 pagesLivestock Science: P. Herold, R. Roessler, A. Willam, H. Momm, A. Valle ZárateNguyen Thi Dieu LinhNo ratings yet
- Multi-Trait Selection Index and Cluster Analyses in Angus CattleDocument8 pagesMulti-Trait Selection Index and Cluster Analyses in Angus CattleClaudia Cristina Paro de PazNo ratings yet
- Genetic Diversity in Farm Animals-A Review-Groeneveld2010Document26 pagesGenetic Diversity in Farm Animals-A Review-Groeneveld2010Raul alejandro Kim gomezNo ratings yet
- Evolutionary Applications - 2021 - Valenza Troubat - Evaluating New Species For Aquaculture A Genomic Dissection of GrowthDocument12 pagesEvolutionary Applications - 2021 - Valenza Troubat - Evaluating New Species For Aquaculture A Genomic Dissection of GrowthRama BangeraNo ratings yet
- C1107021636 PDFDocument21 pagesC1107021636 PDFTeflon SlimNo ratings yet
- Ijisrt20may048 PDFDocument3 pagesIjisrt20may048 PDFKumur JohnNo ratings yet
- Animal Genetics - 2010 - Boettcher - Objectives Criteria and Methods For Using Molecular Genetic Data in Priority SettingDocument14 pagesAnimal Genetics - 2010 - Boettcher - Objectives Criteria and Methods For Using Molecular Genetic Data in Priority SettingSultansarwar SarwarNo ratings yet
- Major Genes and QTL Influencing Wool Production An PDFDocument12 pagesMajor Genes and QTL Influencing Wool Production An PDFSteve Vladimir Acedo LazoNo ratings yet
- MODULE 4 in GeneticsDocument6 pagesMODULE 4 in GeneticsChris TianNo ratings yet
- Green Behavior and DeliveryDocument8 pagesGreen Behavior and DeliveryCao TuấnNo ratings yet
- Kajian Repro Dan PemulDocument11 pagesKajian Repro Dan PemulkusriantynellyNo ratings yet
- PIIS0022030219310884Document16 pagesPIIS0022030219310884Nicolas SierraNo ratings yet
- OhnsonDocument8 pagesOhnsonေကာင္းကင္ တူးမဲ့သူNo ratings yet
- Makalah Genetika DasarDocument7 pagesMakalah Genetika DasarDicky SaputraNo ratings yet
- Iron in Cereal Research PaperDocument6 pagesIron in Cereal Research Paperwkzcoprhf100% (1)
- Mating DesignDocument13 pagesMating DesignShandy Nugraha100% (1)
- Genotype Environment Interaction For Age at First Cal - 2020 - Veterinary and AnDocument5 pagesGenotype Environment Interaction For Age at First Cal - 2020 - Veterinary and Anangelthaiboxyahoo.esNo ratings yet
- Recent Advances in Animal BreedingDocument17 pagesRecent Advances in Animal Breedingdrselvam77No ratings yet
- Functional Genomics and AgricultureDocument16 pagesFunctional Genomics and Agriculturenehadew4No ratings yet
- Genetic Parameters of Traits Associated With The Growth Curve in Segureña SheepDocument8 pagesGenetic Parameters of Traits Associated With The Growth Curve in Segureña Sheepadinda larasNo ratings yet
- n.2205 - Genetics Insights Final ReportDocument129 pagesn.2205 - Genetics Insights Final ReportManilkkaNo ratings yet
- World DevelopmentDocument21 pagesWorld DevelopmentArdiNo ratings yet
- The Impact of Beekeeping On Household Income: Evidence From North-Western EthiopiaDocument8 pagesThe Impact of Beekeeping On Household Income: Evidence From North-Western EthiopiaClaudiu 10134No ratings yet
- Layer Breeding Strategies: An OverviewDocument26 pagesLayer Breeding Strategies: An OverviewSaravanan RamasamyNo ratings yet
- Poultry Production in Nepal: Characteristics, Productivity and ConstraintsDocument6 pagesPoultry Production in Nepal: Characteristics, Productivity and Constraintssabina karkiNo ratings yet
- From The Editors-Animal Selection: The Genomics Revolution: Alessandro Bagnato and Andrea RosatiDocument2 pagesFrom The Editors-Animal Selection: The Genomics Revolution: Alessandro Bagnato and Andrea Rosatiabdelaziz CHELIGHOUMNo ratings yet
- Animal BiotechnologyDocument25 pagesAnimal BiotechnologyFabio Jose GomesNo ratings yet
- Genetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersDocument15 pagesGenetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersIJAR JOURNALNo ratings yet
- Relationships Between Litter Size Sex Ratio and WiDocument28 pagesRelationships Between Litter Size Sex Ratio and WiPeter MukunzaNo ratings yet
- A Future For Transgenic Livestock: John Clark and Bruce WhitelawDocument9 pagesA Future For Transgenic Livestock: John Clark and Bruce WhitelawRoberth Andres LoayzaNo ratings yet
- Aquaculture: Marcelo Augusto Soares Rego, Omar Jorge Sabbag, Roberta Soares, Silvio PeixotoDocument5 pagesAquaculture: Marcelo Augusto Soares Rego, Omar Jorge Sabbag, Roberta Soares, Silvio PeixotoRogerioportelaNo ratings yet
- A Simple Method For On Farm Evaluation of Sheep Welfare - 2022 - Small RuminantDocument7 pagesA Simple Method For On Farm Evaluation of Sheep Welfare - 2022 - Small RuminantBeatrice Geanina RusNo ratings yet
- Genetic Modification and Plant Food Alle PDFDocument9 pagesGenetic Modification and Plant Food Alle PDFIbnu royhanNo ratings yet
- OsvalA MontesinosLpez2023 AmarkerweghtingapproachwihtinfamilygenomicpredictionDocument11 pagesOsvalA MontesinosLpez2023 Amarkerweghtingapproachwihtinfamilygenomicpredictionalanberta26.5No ratings yet
- Agricultural SystemsDocument11 pagesAgricultural SystemsLUIS HUMBERTO MARTINEZNo ratings yet
- PIC Pig DatasetDocument7 pagesPIC Pig DatasetXuân Trường PhạmNo ratings yet
- Contribution of Biological Research To The Development of The Coconut IndustryDocument5 pagesContribution of Biological Research To The Development of The Coconut IndustryPhạm Thị Diệu HuyềnNo ratings yet
- NJAS - Wageningen Journal of Life Sciences: L. TangDocument10 pagesNJAS - Wageningen Journal of Life Sciences: L. TangFathul Hidayat BanureaNo ratings yet
- 24.invited Review Breeding and Ethical Perspectives On Genetically Modified and Genome Edited Cattle - Si PDFDocument17 pages24.invited Review Breeding and Ethical Perspectives On Genetically Modified and Genome Edited Cattle - Si PDFChikorita ChikoritaNo ratings yet
- Multi-Scale Integrated Evaluation of The Sustainability of Large-Scale Use of Alternative Feeds in Salmon AquacultureDocument15 pagesMulti-Scale Integrated Evaluation of The Sustainability of Large-Scale Use of Alternative Feeds in Salmon AquacultureRoger StrandNo ratings yet
- Contribution of The Calving Interval To Dairy Farm Profitability: Results of A Cluster Analysis of FADN Data For A Major Milk Production Area in Southern ItalyDocument12 pagesContribution of The Calving Interval To Dairy Farm Profitability: Results of A Cluster Analysis of FADN Data For A Major Milk Production Area in Southern ItalyRocío ChioNo ratings yet
- Desafios Da Cadeia de Postura 2021Document10 pagesDesafios Da Cadeia de Postura 2021Henrique Carneiro LobatoNo ratings yet
- 1 s2.0 S1751731118001489 MainDocument15 pages1 s2.0 S1751731118001489 MainMaha RanuNo ratings yet
- 482 2585 1 PBDocument4 pages482 2585 1 PBali97lol97No ratings yet
- Journal Homepage: - : IntroductionDocument9 pagesJournal Homepage: - : IntroductionIJAR JOURNALNo ratings yet
- Assignment On Genetics and Agriculture by AkhilaDocument26 pagesAssignment On Genetics and Agriculture by AkhilaAkhila ManuNo ratings yet
- Swine Production, Productivity and Breeding Practices in EthiopiaDocument10 pagesSwine Production, Productivity and Breeding Practices in EthiopiaDiriba WakgariNo ratings yet
- Title: Revised Investigator: Institution: Co-Investigators: Date SubmittedDocument23 pagesTitle: Revised Investigator: Institution: Co-Investigators: Date SubmittedWella ChristafariNo ratings yet
- Herrero Etal 2017 Farming and The Geography of Nutrient Production - LANPLHDocument11 pagesHerrero Etal 2017 Farming and The Geography of Nutrient Production - LANPLHVictor BonillaNo ratings yet
- Fostering Innovation for Agriculture 4.0: A Comprehensive Plant Germplasm SystemFrom EverandFostering Innovation for Agriculture 4.0: A Comprehensive Plant Germplasm SystemNo ratings yet
- 174 468 1 PBDocument9 pages174 468 1 PBAlfin Nur FauziNo ratings yet
- Dairy Cows Welfare Quality in Tie-Stall Housing System With or Without Access To ExerciseDocument11 pagesDairy Cows Welfare Quality in Tie-Stall Housing System With or Without Access To ExerciseLilisNo ratings yet
- Genomic Breed Prediction in New Zealand Sheep: Researcharticle Open AccessDocument15 pagesGenomic Breed Prediction in New Zealand Sheep: Researcharticle Open AccessAlfin Nur FauziNo ratings yet
- Perennial ryegrass breeding in NZ for dairy industryDocument22 pagesPerennial ryegrass breeding in NZ for dairy industryAlfin Nur FauziNo ratings yet
- Han2013 Article GeneticVariationsInTheMyostatiDocument6 pagesHan2013 Article GeneticVariationsInTheMyostatiAlfin Nur FauziNo ratings yet
- Application of New Technologies in Sheep BreedingDocument15 pagesApplication of New Technologies in Sheep BreedingAlfin Nur FauziNo ratings yet
- Lodging EstablishmentsDocument2 pagesLodging EstablishmentsMich A. Gonzales0% (1)
- 20090716FoxReiki PDFDocument40 pages20090716FoxReiki PDFAgustina RomeroNo ratings yet
- Cast Resin Planning Guidelines GEAFOL PDFDocument24 pagesCast Resin Planning Guidelines GEAFOL PDFtenk_man100% (1)
- M 200Document3 pagesM 200Rafael Capunpon VallejosNo ratings yet
- Smart medical system monitors dementia patients' medicationDocument9 pagesSmart medical system monitors dementia patients' medicationKresnaNo ratings yet
- Chapter 43 - Lead - 2015 - Handbook On The Toxicology of MetalsDocument57 pagesChapter 43 - Lead - 2015 - Handbook On The Toxicology of MetalsChanWingSanNo ratings yet
- Name: Sport: Movement PrepDocument24 pagesName: Sport: Movement PrepS HNo ratings yet
- Animals Pets Board GameDocument1 pageAnimals Pets Board GameOlga ZasNo ratings yet
- Online Medicine Industry Competitor AnalysisDocument14 pagesOnline Medicine Industry Competitor Analysispawangadiya1210No ratings yet
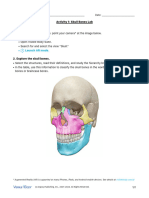
- Lab Manual - Skull Bones - English - Student - Fill inDocument6 pagesLab Manual - Skull Bones - English - Student - Fill inthejaskumar2026No ratings yet
- Ventilation System Comparison - Constant Air Volume (CAV) and Variable Air Volume (VAV)Document15 pagesVentilation System Comparison - Constant Air Volume (CAV) and Variable Air Volume (VAV)ankurNo ratings yet
- BC-5300 Preventive Maintenance Kit ManualDocument8 pagesBC-5300 Preventive Maintenance Kit ManualAbdel MUNDENNo ratings yet
- Caffeine Natural Coffee BeanDocument1 pageCaffeine Natural Coffee BeanMayang TariNo ratings yet
- Wartsila Tribo PackDocument7 pagesWartsila Tribo Packsuper_seeker100% (1)
- Jear C PB 2018 PDFDocument174 pagesJear C PB 2018 PDFArasu RajendranNo ratings yet
- Beginning Algebra 9th Edition Tobey Solutions Manual 1Document39 pagesBeginning Algebra 9th Edition Tobey Solutions Manual 1kyle100% (30)
- Reactor & Impeller Design in Hydrogenation: GBHE Technical Bulletin CTB #79Document13 pagesReactor & Impeller Design in Hydrogenation: GBHE Technical Bulletin CTB #79manuNo ratings yet
- Telephone Directory Updated 24.04.2020Document139 pagesTelephone Directory Updated 24.04.2020Manpal SinghNo ratings yet
- Nikola Tesla Would Have Celebrated Christmas On January 7thDocument3 pagesNikola Tesla Would Have Celebrated Christmas On January 7thAnonymous DfeajF100% (1)
- DBXLH 8585B VTMDocument4 pagesDBXLH 8585B VTMmau_mmx5738No ratings yet
- Understanding Immunomodulatory DrugsDocument4 pagesUnderstanding Immunomodulatory DrugsMark Russel Sean LealNo ratings yet
- RMO NO. 66-2016 - DigestDocument2 pagesRMO NO. 66-2016 - DigestJuliannaMendozaMaleNo ratings yet