Professional Documents
Culture Documents
Jurnal Mitogenic
Uploaded by
Haykal Estu BhismoroOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Jurnal Mitogenic
Uploaded by
Haykal Estu BhismoroCopyright:
Available Formats
Am J Physiol Regulatory Integrative Comp Physiol
278: R1651–R1660, 2000.
Mitogenic whey extract stimulates wound repair activity
in vitro and promotes healing of rat incisional wounds
TIMOTHY E. RAYNER,1 ALLISON J. COWIN,1 J. GRAY ROBERTSON,1
RODNEY D. COOTER,2 RICHARD C. HARRIES,2 GEOFFREY O. REGESTER,1
GEOFFREY W. SMITHERS,3 CHRIS GODDARD,4 AND DAVID A. BELFORD4
1Cooperative Research Centre for Tissue Growth and Repair, Child Health Research Institute,
Women’s and Children’s Hospital, North Adelaide 5006; 2Plastic and Reconstructive Surgery,
University of Adelaide, Department of Surgery, The Queen Elizabeth Hospital, Woodville 5011;
4Commonwealth Scientific and Industrial Research Organization Division of Human Nutrition,
Adelaide BC 5000; and 3Food Science Australia, Highett, Victoria 3190, Australia
Rayner, Timothy E., Allison J. Cowin, J. Gray Robert- growth factor (TGF)-b (34, 38, 39), platelet-derived
son, Rodney D. Cooter, Richard C. Harries, Geoffrey O. growth factor (PDGF) (17, 18, 39), and epidermal
Regester, Geoffrey W. Smithers, Chris Goddard, and growth factor (EGF) (19) families, the interleukins (26),
David A. Belford. Mitogenic whey extract stimulates wound and colony stimulating factor (24). Importantly, these
repair activity in vitro and promotes healing of rat incisional factors have been shown to reverse the healing deficit
wounds. Am J Physiol Regulatory Integrative Comp Physiol
associated with diabetes (17), steroid administration
278: R1651–R1660, 2000.—The ability of single growth fac-
tors to promote healing of normal and compromised wounds (38), cytotoxic therapy (25), infection (24), radiation (9,
has been well described, but wound healing is a process 35), and ischemia (49). However, the activity of these
requiring the coordinated action of multiple growth factors. factors in promoting wound repair on the experimental
Only the synergistic effect on wound healing of combinations animal has not been reproduced in clinical studies of
containing at most two individual growth factors has been chronic wound healing (43). Whereas there are several
reported. We sought to assess the ability of a novel milk- possible reasons for this discrepancy, it now seems
derived growth factor-enriched preparation [mitogenic bovine likely that the proinflammatory environment of the
whey extract (MBWE)], which contains six known growth chronic wound adversely affects the stability and activ-
factors, to promote repair processes in organotypic in vitro ity of the applied growth factors (29, 50). Thus the
models and incisional wounds in vivo. MBWE stimulated the ability of any single factor to enhance the multiple
contraction of fibroblast-populated collagen lattices in a dose-
aspects of wound repair to produce a clinically signifi-
dependent fashion and promoted the closure of excisional
wounds in embryonic day 17 fetal rat skin. Application of cant outcome may be limited.
MBWE increased incisional wound strength in normal ani- Several studies have now clearly shown that wound
mals on days 3, 5, 7, and 10 and reversed the decrease in repair is the result of the temporally coordinated local
wound strength observed following steroid treatment. Wound expression of multiple families of growth factors and
histology showed increased fibroblast numbers in wounds their receptors (5). It would therefore seem reasonable
from normal and steroid-compromised animals. These data to hypothesize that the application of mixtures of
suggest the mixture of factors present in bovine milk exerts a growth factors to the healing wound may be more
direct action on the cells of cutaneous wound repair to effective than the addition of a single factor. Indeed,
enhance both normal and compromised healing. several studies have shown this to be the case. In
growth factor; wound healing; skin; fetal wound healing; particular, PDGF interacts with IGF-I (27), IGF-II (16),
fibroblast-populated collagen lattice EGF, and TGF-b (25), insulin (18), and TGF-a (7) to
promote the repair of a variety of wounds. EGF and
insulin act synergistically to promote collagen synthe-
sis in the diabetic rat (19). Similarly, coapplication of
THE TOPICAL APPLICATION OF several classes of growth IGF-I and its binding proteins is more effective at
factors has been shown to enhance all aspects of wound promoting wound repair than application of IGF-I
repair, including inflammation, matrix synthesis, wound alone (22, 33).
contraction, epithelialization, and tissue remodelling Bovine milk is a rich source of several classes of
(6). Studies to date have largely focussed on the effects growth factors, including PDGF (46), IGF-I and -II (11),
of single recombinant growth factors, including mem- and TGF-b (21). We have reported that cation-exchange
bers of the fibroblast growth factor (FGF) (17, 30), fractionation of bovine whey results in a 100- to 200-
insulin-like growth factor (IGF) (22, 48), transforming fold enrichment of the growth factor activity present in
milk (10). The resultant fraction, mitogenic bovine
whey extract (MBWE), was shown to stimulate the
The costs of publication of this article were defrayed in part by the
payment of page charges. The article must therefore be hereby
growth of cells of mesodermal origin, although, due to
marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 its TGF-b content, was found to be inhibitory to the
solely to indicate this fact. division of epithelial cells (3, 45). This fraction also
http://www.ajpregu.org 0363-6119/00 $5.00 Copyright r 2000 the American Physiological Society R1651
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
R1652 MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR
contains significant concentrations of IGF-I and -II, sured (Limulus Amebocyte Lysate assay, BioWhittaker, Walk-
IGF binding proteins 2 and 3, PDGF, and FGF-1 and -2, ersville, MD).
although together these known growth factors account
Fibroblast-Populated Collagen Lattice Contraction
for only 50% of growth activity present (2, 44). Maximal
growth of fibroblast lines in response to MBWE signifi- Human diploid SF were maintained in DMEM supple-
cantly exceeded that in the presence of several indi- mented with penicillin (60 mg/l), streptomycin (100 mg/l),
vidual recombinant factors including PDGF, IGF-I, fungizone (1 mg/l), and 10% FBS. Stock cultures were main-
TGF-b, FGF-2, or EGF (3). tained in 75-cm2 flasks (Corning, NY) in 5% CO2-95% air at
The aim of the current study was to investigate the 37°C and routinely passaged after suspension in trypsin
(0.125%)/EDTA (0.5 mM) in Dulbecco’s phosphate buffered
wound repair activity of the milk-derived growth factor saline.
mixture. In vitro organotypic models initially were A solution of rat tail collagen (2 mg/ml) was mixed with an
used to characterize the tissue repair activity of MBWE, equal volume of 23 DMEM at 4°C before addition of human
and an in vivo model of incisional wound healing was SFs (200,000 cells/ml final concentration) and [3H]inulin (to
used to assess the ability of MBWE to promote cutane- 30,000–40,000 counts · min21 · ml21). Five-hundred microli-
ous repair in normal animals and reverse the deficit in ters of this solution were poured into each well of a 48-place
healing associated with steroid administration. tissue culture plate (Costar, Cambridge, MA) and allowed to
set by incubation at 37°C for 30 min. Gels were freed from the
surrounding tissue culture plastic by rimming the margin
MATERIALS AND METHODS
with a 25-gauge needle. Dilutions of MBWE were applied to
Materials the top of the gels, which were then maintained in a humidi-
fied atmosphere of 5%CO2-95% air at 37°C. Control wells
Human diploid skin fibroblasts (SF) were provided by the containing either serum-free or FBS-supplemented medium
Women’s and Children’s Hospital (North Adelaide, Australia) were incorporated on each plate. Fibroblast-induced contrac-
and DMEM obtained from Flow Laboratories (Irvine, Scot- tion was assessed by counting the radioactivity remaining in
land). Fetal bovine serum (FBS) was obtained from Cytosys- the gel after 24-h culture. Data were fit to a four-parameter
tems (Castle Hill, Australia) and [3H]inulin (1.18 Ci/mmol) equation with the aid of a nonlinear curve-fitting program
from Amersham (Buckinghamshire, UK). (Tablecurve, Jandel Scientific, San Rafael, CA).
Male Sprague-Dawley rats (250–300 g) were purchased
from the University of Adelaide. All experiments were ap- Fetal Skin Organ Culture
proved by the appropriate institutional animal care and
Wounding and culture of fetal rat skin were undertaken as
ethics committee of the Women’s and Children’s Hospital, The
previously described (1). Pregnant rats were killed by CO2
Queen Elizabeth Hospital, or CSIRO Human Nutrition follow-
asphyxiation, the fetuses (E16–E17) removed from the uterus,
ing the Australian Code of Practice for the Care and Use of
weighed, and placed in Hank’s balanced salt solution. A 1 3
Animals for Scientific Purposes. Methylprednisolone (Depo-
1-cm piece of skin was dissected from the back of each fetus
Medrol, 20 mg/ml) was purchased from Upjohn (Rydalmere,
and a 1-mm diameter wound created in each explant using a
New South Wales, Australia). Fluothane anesthetic (Halo-
squared-off, sharpened 19-gauge needle. The wound margin
thane) was obtained from ICI Australia (Victoria, Australia)
was marked by dipping the cutting needle into Indian ink
and used in combination with oxygen and nitrous oxide (BOC
immediately before wounding. The wounded skin was then
Gases, South Australia, Australia). Collagen was extracted
mounted on a six-pin cradle and placed in a 12-well tissue-
from rat tail tendons in a solution of 0.1% acetic acid
culture dish (Costar). After three washes with medium,
according to the method of Bell et al. (4). Before use as a
serum-free DMEM or DMEM/MBWE (2.5 mg/ml) was added
vehicle matrix, the collagen was diluted in sterile water to a
in a final volume of 2 ml. Wounds were photographed using a
final concentration of 1 mg/ml and brought to neutrality with
standard focal length jig at the time of culture and at 24, 48,
phosphate-buffered saline and NaOH.
and 72 h. Wound area was calculated by tracing the wound
margin onto acetate sheets which were then scanned into a
Production of MBWE computer (Apple Macintosh Onscanner connected to an Apple
Macintosh LCIII) equipped with an image analysis program
Fractionation of whey was carried out as previously de-
(Prismview, Dapple Systems, Sunnyvale, CA). For histology,
scribed (10). Briefly, pasteurized whey obtained as a byprod-
cultures were fixed in methacarn for 2 h before storage in 70%
uct of cheese manufacture was clarified by passage through a
alcohol and processing by graded dehydration. Skin pieces
0.8-µm ceramic filter (Membralox, Bajet, France). The ultra-
were embedded vertically in wax blocks and 3-µm sections
filtrate was adjusted to pH 6.5 and applied to a 30-cm
taken through the wound. Sections were stained with hema-
diameter column (Moduline, Beverly, MA) packed with 5 l of
toxylin and eosin, viewed, and photographed.
S-Sepharose Fast Flow cation-exchange resin (AMRAD Phar-
macia Biotech, Victoria, Australia) equilibrated with 50 mM Incisional Wound Healing Model
sodium citrate buffer at pH 6.5. After washing the column
with the same buffer, the absorbed material was eluted with Ability to promote repair of an incisional wound was
0.5 M NaCl. This eluate was desalted, concentrated, and investigated as previously described (34, 38). After induction
freeze dried. Growth activity was confirmed using a dye- of general anesthesia by inhalation of halothane, paired
binding cell growth assay as previously described (3). TGF-b 6-cm incisions were created through the panniculus carnosis
concentration was determined using an Mv1Lu cell bioassay either side of the midline on the dorsal surface of the rat using
(45). IGF-I concentration was measured by radioimmunoas- a standard template. MBWE was dissolved in the collagen
say after removal of IGF binding proteins by acid gel- vehicle at the indicated concentrations and a single 200-µl
permeation HPLC (37). MBWE was screened for bovine dose of this formulation applied to one wound. The same
diarrhea virus, infectious bovine rhinotracheitis, parainflu- volume of collagen vehicle was applied to the contralateral
enza type 3 and coliforms. Endotoxin levels were also mea- wound. The collagen was noted to cover the wound surface
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR R1653
and gelled within 1–2 min. Left- and right-sided wounds were present in MBWE, including PDGF and FGF, were not
randomized with respect to treatment and were closed using routinely determined. In addition, MBWE was shown
five sutures (4/O silk, Dyneck, South Australia, Australia). to be free of all microbiological contaminants tested and
Care was taken during suturing to evert the wound margins contained ,1 ng/ml endotoxin.
to ensure dermal-dermal contact along the length of the
wound. All rats were housed individually after surgery and Effect of MBWE on Fibroblast-Induced Gel Contraction
were weighed the day after surgery and then every 2 days. At
various time points after wounding, rats were killed by CO2 The ability of MBWE to stimulate growth of several
asphyxiation and the pelts carefully removed. A standard fibroblast cell lines (3) led us to further examine its
template was used to cut four strips of 0.5-cm width through repair activity using organotypic models of wound
each wound (i.e., 1 strip through each of the wound segments healing. MBWE markedly stimulated fibroblast-in-
bordered by two sutures, excluding the top and bottom wound duced gel contraction in a dose-responsive manner
margins) and either peak breaking strength measured using
when applied to fibroblast-populated collagen lattices
a M1000E (Mecmesin, West Sussex, UK) tensiometer (time-
course study) or a breaking profile of each strip generated (FPCL; Fig. 1). The dose that exerts half-maximal effect
using a PCM2500EL tensiometer (Mecmesin) connected to a above baseline in this assay over a series of similar
personal computer (dose-response study). The latter allowed experiments was 195 6 44 µg/ml (mean 6 SE; n 5 3).
calculation of peak load, yield load (defined as the point at
which the gradient change in load over a set displacement Effect of MBWE on Wound Closure in Organ Cultured
interval becomes ,20% of the calculated start gradient), force Fetal Rat Skin
absorbed to yield load, and force absorbed to peak load.
The effect of MBWE on wound repair was further
Histology examined using a fetal skin organ culture model. Skin
harvested from E16–E17 rat embryos retains the abil-
Samples of MBWE and collagen-treated wounds were ity to heal an excisional wound in serum-supplemented
collected and placed in 10% Formalin fixative immediately suspension culture (1) and allows the effect of agents
after harvesting and processed by graded dehydration. Three with potential wound-healing activity to be evaluated
sections (3 µm) were cut through each wound, stained with in an organ culture model isolated from blood-borne or
hematoxylin and eosin, and scored by two independent observ-
environmental factors. Wounded E16–17 rat skin cul-
ers (blinded). Each section was scored for mononuclear cell
and fibroblast infiltration. Scores of zero to four were recorded tures were established as described and cultured in
for each section, and the average of three sections from each DMEM alone or DMEM plus MBWE (2.5 mg/ml). In
scorer was used to obtain an average score for the wound, DMEM alone, wounds contracted to 35–40% of the
which was used to calculate overall group means. original wounded area over 72 h, although they never
completely closed (Fig. 2A, a and c) (1). Addition of
Data Analysis MBWE to the medium promoted complete closure over
Five percent of strips in the steroid-treated group showed
evidence of hemorrhage or dehiscence before dissection and
were removed from the wound strength data set. Similarly,
spontaneous separation of the dermal margins of the wound
during processing for tensiometry (i.e., once they had been
removed from the support of the sutures) was recorded as
dehiscence, and the strip was excluded from wound-strength
data analysis. All results are presented as means 6 SE with
wound-strength data analyzed using Student’s t-test (dose-
response study) or paired t-test (time-course study). Histology
scores were analyzed by Kruskall-Wallis one-way analysis of
variance on ranks with multiple comparisons versus the
control group (combined data) performed using Dunn’s
method. P , 0.05 was considered significant.
RESULTS
Activity of MBWE
Cell growth activity of MBWE was confirmed using
Balb/c 3T3 fibroblasts and SFs as previously described
(3). Growth activity corresponding to that observed in Fig. 1. Fibroblast populated collagen lattice (FPCL) contraction.
10% FBS for Balb/c 3T3 fibroblasts and SFs was FPCLs were prepared such that each 500 µl gel contained 1 mg/ml
collagen plus 105 skin fibroblasts and [3H]inulin [30–40,000 counts/
achieved with 150 and 610 µg/ml MBWE, respectively. min (cpm)]. Gels were freed from side of well and cultured in a
The batch of MBWE used in the current study con- dilution series of mitogenic bovine whey extract (MBWE). Control
tained 1.6 ng/mg active TGF-b and 61 ng/mg total wells containing either serum-free or fetal bovine serum (FBS)-
TGF-b bioactivity as determined by Mv1Lu cell bioas- supplemented medium were incorporated on each plate. At 24 h,
culture gels were placed in scintillation fluid and the radioactivity
say before and after acid activation, respectively. The remaining in gel determined in a b-counter. Points represent means 6
batch of MBWE also contained 32 ng/mg IGF-I. The SE of triplicate gels. Control values were: serum-free DMEM 23,140 6
concentrations of other growth factors known to be 840 cpm; DMEM/10% FBS 7,990 6 640 cpm (means 6 SE, n 5 4).
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
R1654 MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR
epidermal layer over the wound edge to form a ‘‘plug’’ of
epithelium between the dermal margins (Fig. 3B).
Time Course of Response to MBWE
To examine the ability of MBWE to stimulate wound
healing in normal rats and determine the time course of
this response, MBWE was formulated at 2.5 mg/ml,
corresponding to a total dose of 0.5 mg/wound, and
applied to linear incisions on the dorsum of adult rats.
This concentration of MBWE has previously been shown
to stimulate maximal growth of human SFs in vitro (3),
although concentrations up to 5 mg/ml were tolerated
by these cells without a significant decrease in the
maximal growth response. Wound strength was as-
sessed at days 3, 5, 7, 10, 14, 21, and 42 after wounding.
The peak load tolerated by MBWE- and vehicle-
treated wounds is shown in Table 1. Highly significant
increases in wound strength in response to topical
MBWE were seen on days 5, 7, and 10, corresponding to
average increases of 44%, 28%, and 20% over control
wounds, respectively. The strength of MBWE-treated
wounds on days 3 and 21 were increased an average of
21% and 10% over control values, respectively. Break-
Fig. 2. Fetal excisional wound closure. Fetal skin was dissected from
dorsum of E17 rats and wounded using a squared off 19-gauge needle.
Wound margin was marked by dipping cutting needle into Indian ink
immediately before wounding. A: wounds are shown at time of
culture (a and b) and after 72 h in serum-free DMEM (c) or DMEM 1
2.5 mg/ml MBWE (d ). B: contraction curves from triplicate experi-
ments after measurement of wound areas at times shown for
wounded skin incubated in serum-free DMEM (s) or DMEM 1 2.5
mg/ml MBWE (r).
the 72-h culture period (Fig. 2A, b and d). The contrac-
tion of wounds in skin explants treated with MBWE
was greatly accelerated compared with skin incubated
in DMEM alone (Fig. 2B). The wound areas of MBWE-
treated explants at 24 and 48 h were 20 6 10% and
2.8 6 2.2% their original area, respectively (Fig. 2B,
means 6 SE of triplicate cultures). In agreement with a Fig. 3. Histology of excisional wounds in fetal rat skin after 48 h
previous study (1), histology through the wound after culture in MBWE. Experimental details are given in Fig. 2 legend. At
48 h in DMEM alone showed no evidence of epithelial or 48 h culture, skin explants were fixed in methacarn, processed for
mesenchymal movement into the wound defect (Fig. wax embedding, and 3-µm sections were taken through wound that
was identified by concentration of ink particles. Sections were stained
3A). In contrast, histology through healed wounds after with hematoxylin and eosin. A: wound margin after 48 h in serum-
48 h in the presence of MBWE showed that final wound free DMEM. B: healed wound after 48 h in DMEM 1 2.5 mg/ml
closure had been achieved by the movement of the MBWE.
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR R1655
Table 1. Effect of MBWE on wound strength tensiometer. This degree of wound breakdown in the
in normal rats steroid-treated group was reduced to 13% in wounds
treated with 0.5 mg/wound MBWE, whereas the dehis-
Breaking Strength, g cence rate in wounds treated with 0.25 and 2.0 mg/
Day Collagen MBWE P wound MBWE was consistent at 21%. Wounds treated
with the 0.5 mg/wound dose of MBWE also exhibited a
3 27.8 6 2.5 33.5 6 2.4 0.02
5 45.4 6 3.6 65.4 6 4.8 0.0003 significant increase in both the force absorbed to yield
7 85.5 6 6.5 109.4 6 6.0 0.0003 load (not shown) and force absorbed to peak load
10 169.8 6 10.8 203.5 6 10.6 0.0003 compared with vehicle-treated wounds (Fig. 4B). In-
14 277.3 6 16 297.4 6 13.8 0.10 deed, 0.5 mg/wound MBWE increased the wound
21 958.6 6 50.1 1,053 6 59.2 0.02
42 3,300 6 110 3,289 6 80.4 0.9
strength to levels recorded from normal control wounds
Values are means 6 SE for 16 wounds (64 strips) pooled from 2
separate experiments. Paired linear incisions were created on the
dorsal surface of adult male rats as described in MATERIALS AND
METHODS. Mitogenic bovine whey extract (MBWE; 2.5 mg/ml of
collagen) was applied to one wound and collagen vehicle was only
applied to the contralateral wound. On the indicated days after
wounding 1 group was killed and the maximal load (grams) tolerated
by 4 strips of 0.5-cm width were taken through each wound deter-
mined using a tensiometer. Data were analyzed by paired t-test.
ing strengths on days 14 and 42 were not significantly
different from vehicle-treated controls.
Dose Response to MBWE
A series of studies aimed at determining the optimal
dose and assessing the histological picture of the
MBWE-treated wounds was undertaken using both
normal and steroid-compromised animals. Steroid-
treated rats received an intramuscular injection of
methylprednisolone (15 mg/kg) immediately before sur-
gery. MBWE was formulated at 10, 2.5, and 1.25 mg/ml,
each wound therefore receiving a total of 2, 0.5, and
0.25 mg of MBWE, respectively. As the maximal re-
sponse to MBWE in normal animals occurred 5 days
after wounding and application of the extract, all
wounds were harvested at this time point.
Normal animals. The force absorbed by each strip
harvested from normal animals 5 days after wounding
is shown in Fig. 4A. Both force absorbed to yield load
(not shown) and force absorbed to peak load were
significantly increased in wounds treated with 0.5
mg/wound MBWE compared with vehicle-treated
wounds. In addition, the yield load and peak load in the
MBWE-treated wounds were increased an average of
62% and 25% over control wounds, respectively (data
not shown). Application of either 0.25 or 2.0 mg/wound
MBWE did not significantly alter wound strength.
Steroid-treated animals. Administration of methyl-
prednisolone on the day of wounding induced an aver- Fig. 4. Wound strength in normal (A) and steroid-treated animals (B)
age 15% weight loss compared with an 8% weight gain in response to MBWE. Paired linear incisions were created on the
in the normal group over the course of the experiment dorsal surface of adult male rats. MBWE was applied to one wound at
and significantly reduced all indices of wound strength. indicated doses and collagen vehicle was only applied to the contralat-
Yield load, peak load, force absorbed to yield load, and eral wound. Animals in the steroid group were given an intramuscu-
lar injection of 15 mg/kg methylprednisolone at time of surgery.
force absorbed to peak load of vehicle-treated wounds Animals were killed 5 days after wounding and 4 strips of 0.5-cm
were decreased an average of 44%, 39%, 42%, and 39%, width were taken through each wound and placed in a tensiometer.
respectively, in the steroid animals (P , 0.0001 for all). Force absorbed to peak load was calculated from load-displacement
Whereas only ,1–2% of all strips taken through wounds curves. Bars represent means 6 SE of indicated number of strips (in
parentheses) pooled from 2 experiments for both MBWE (filled bars)-
on normal animals showed any signs of dehiscence, and matched vehicle-treated (open bars) wounds. Different number of
28% of strips cut through vehicle-treated wounds on strips analyzed reflects incidence of wound strip dehiscence in
steroid-treated rats disrupted before placement in the respective treatment groups. * P , 0.05 vs. vehicle-treated wounds.
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
R1656 MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR
(Fig. 4A). Furthermore, the values for both the yield in Fig. 6. Comparison of Fig. 5, A and C, shows the
load and peak load were increased an average of 60% difference in wound histology expected between normal
above their vehicle-treated controls (data not shown). and steroid-treated rats, which is characterized by the
As observed with wounds from normal animals, applica- marked absence of cellular infiltrate and granulation
tion of either 0.25 or 2.0 mg of MBWE did not signifi- tissue. Administration of methylprednisolone signifi-
cantly affect the force absorbed by the wound compared cantly reduced the scores, reflecting fibroblast and
with their vehicle-treated controls (Fig. 4B). inflammatory cell infiltration by 46% and 34%, respec-
Wound histology. Representative sections taken tively (Fig. 6, A and B). Treatment with MBWE in-
through wound samples from normal and steroid rats creased the cellular infiltrate in wounds on both normal
are shown in Fig. 5 with semiquantitative histology and steroid-treated rats (Fig. 5, B and D). This response
scores of sections from all treatment groups presented to MBWE was most noticeable in sections from wounds
Fig. 5. Wound histology. Photomicro-
graphs show cross sections through day
5 incisional wounds from normal (A and
B) and steroid-treated (C and D) rats
that received MBWE (B and D) or colla-
gen vehicle only (A and C) at time of
wounding. Sections were stained with
Masson’s trichrome and photographed
at 3250 magnification.
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR R1657
with either 0.25 or 0.5 mg/wound MBWE showed a
significant increase in fibroblast score. MBWE had no
significant effect on the inflammatory score in wounds
on both normal and steroid-treated animals (Fig. 6B).
DISCUSSION
We previously showed that cation-exchange chroma-
tography results in a 100- to 200-fold enrichment of
the growth factor component of bovine milk. Although
consisting of only 0.5% of whey protein, it contains the
bulk of growth factor activity present in bovine milk
(10). Importantly, this fraction contains several growth
factors and their binding proteins (2) that have been
shown by others to act in synergistic fashion in the
wound, including IGF-I and PDGF (27), IGF-II and
PDGF (16), and IGF-I and IGF binding protein 3 (33).
In addition, MBWE contains significant concentrations
of members of the TGF-b family, with TGF-b1, a factor
known to significantly enhance the healing strength
of an incisional wound (34). Approximately 85% of the
TGF-b activity in MBWE is accounted for by TGF-b2,
which exists as an acid labile 80-kDa species consistent
with the small latent complex of TGF-b (45). In this
regard, it is interesting to note that some 30–40% of
platelet-derived TGF-b is retained in the clot as
the small latent complex, where it may act as a slow
release form of the molecule during the wound repair
process (15).
The activity of the growth factor-enriched milk-
derived fraction in promoting different aspects of tissue
repair was investigated in the first instance using in
vitro organotypic models. The ability of fibroblasts
incorporated in a collagen gel to reorganize the collagen
fibers and cause gel contraction is considered the in
vitro counterpart of wound contraction in the adult
mammal (4, 32). This phenomenon has been found to be
dependent on the trophic activity of FBS (14). However,
in the present study, MBWE was able to substitute for
serum in stimulating fibroblast-mediated collagen lat-
tice contraction (Fig. 1). MBWE-induced contraction
was dose dependent, with the maximal effect over 24 h
found to be at least equivalent to that observed in
serum-supplemented culture (not shown). Given that
Fig. 6. Semiquantitative assessment of wound histology. Sections
through wounds on both normal (open bars) and steroid-treated
the individual growth factors EGF, PDGF, and basic
(filled bars) rats were scored for degree of fibroblast (A) or inflamma- FGF (bFGF) failed to stimulate FPCL contraction (32),
tory cell infiltration (B). Wounds received indicated dose of MBWE or it would appear that the novel mixture of growth
collagen vehicle only at time of wounding. Data show mean score for factors contained within MBWE may account for the
each treatment group (6 SE), which was derived from triplicate response observed in this assay, with IGF-I or TGF-b
sections through each wound sample with number of wounds as-
sessed in each group shown in parentheses. * P , 0.05 vs. vehicle- likely to be at least partly responsible for the activity
treated wounds (combined across all groups). evident in MBWE (13, 41).
The assertion that a mixture of individual growth
factors such as that present in MBWE may be effective
on steroid-treated animals, the degree of wound cellu- at stimulating tissue repair is further supported by the
larity comparable with sections from normal control ability of MBWE to promote wound closure in organ-
wounds (compare Fig. 5, A and D). This increase in cultured fetal skin. Skin harvested from the E16–17
cellularity was due to an influx of fibroblasts into the embryo is able to heal a wound in vitro by a combined
wound, which was not accompanied by a heightened process of mesenchymal contraction and substrate-
inflammatory response (Fig. 6, A and B). Within the independent movement of the epithelium over the
steroid-treated group, all doses of MBWE significantly dermal margins of the wound to effect final closure (1).
increased fibroblast numbers compared with their ve- This repair response is dependent on the presence of
hicle-treated controls. In normal rats, wounds treated serum but is not promoted by the addition of single
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
R1658 MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR
recombinant factors such as IGF-I, EGF, bFGF, TGF-b, bimodal effect of TGF-b and bFGF on the volume of new
or PDGF. Although wound closure is likely to result granulation tissue detected in rabbit ear excisional
from the coordinated action of epithelial cells to mobi- wounds has also been reported (40).
lize contractile proteins and close the wound by a The possible complex interactions among the known
purse-string mechanism (28), the trophic requirement growth factors in MBWE render any discussion of its
for this form of epithelial repair is unknown. MBWE mechanism of action in the wound somewhat specula-
was able to substitute for serum in promoting wound tive. Furthermore, it is impossible to rule out effects on
closure, histology clearly revealing the presence of wound healing by components of MBWE that are not
epithelium spanning the dermal margins of the wound growth factors. Moreover, although the activity of
(Fig. 3). Nevertheless, the dermal margins were consis- MBWE in steroid-treated rats would suggest a direct
tently more approximated after culture in MBWE than action on the wound, we cannot exclude indirect effects
in serum-supplemented cultures compared with previ- of the components of MBWE on the cells of wound
ously published results (1), possibly reflecting the ac- repair. Nevertheless, comparative studies using single
tion of the factors in MBWE on the mesenchymal layer growth factors may provide some clues. PDGF, TGF-b,
to enhance wound contraction. Thus the mixture of and FGF all stimulate granulation tissue deposition.
factors contained in MBWE has the capacity to induce TGF-b1 in particular has been shown to act directly on
novel effects on fetal wound closure that are not ob- the cells of cutaneous healing to induce the fibrotic and
served after application of individual factors, suggest- angiogenic components of repair in vivo (42). Thus
ing that appropriate combinations of growth factors TGF-b1 is effective in promoting repair of wounds on
may act in a coordinated fashion to stimulate the animals rendered monocytopenic by steroid administra-
sequence of events required for successful healing. We tion (38) or total body irradiation (9), although it is
cannot, however, rule out the possibility that an as yet ineffective in rats whose dermal fibroblasts have been
unidentified molecule(s) may be present in MBWE that damaged by surface irradiation (9). In contrast, PDGF
has unique and singular effects on the repair process. does not directly stimulate collagen synthesis (42) and
All surface wounds heal by a process of wound cannot reverse the wound healing deficit associated
contraction, matrix synthesis and remodelling, and with circulating and wound monocytopenia as a result
epithelialization. Apposition of the wound margins of steroid treatment (38), although it was able to
allows healing by primary intention, which is mainly improve wound repair in animals whose dermal fibro-
achieved by matrix synthesis. Thus the ability of a blasts had been damaged by surface irradiation by
topical agent to enhance the strength of an incisional recruiting macrophages to the wound (35).
wound reflects a direct or indirect ability to enhance The degree of improvement in wound strength ob-
wound fibroblast division, migration into the wound, or served in response to MBWE 5 days after wounding
biosynthetic capacity. The ability of MBWE to promote steroid-treated rats is greater than that reported follow-
wound repair was most evident in steroid-treated ani- ing administration of recombinant TGF-b1 in a compa-
mals, suggesting a direct action of the growth factors in rable study (38), suggesting the potential synergistic
MBWE on the cells of wound repair. Steroid administra- action of factors present in MBWE. In this regard,
tion profoundly inhibits both the inflammatory re- IGF-I and -II are also known to promote fibroblast
sponse to wounding and the expression of procollagen division (8), protein, collagen, and fibronectin synthesis
genes by SFs (12). A single dose of methylprednisolone (20), and local administration of IGF-I has been shown
at or before wounding inhibits wound contraction (23), to reverse the effects of steroids on inhibition of granu-
synthesis of granulation tissue, and angiogenesis and lation tissue ingrowth to a subcutaneous chamber (48).
decreases wound strength (38). In the current study, all Coadministration of IGF-I with its binding proteins
indices of wound strength were reduced by ,40% of increases wound strength (22). Similarly, both acidic
control values 5 days after administration of methyl- and basic FGF have been shown to increase wound
prednisolone. The most effective dose of MBWE (0.5 strength (30, 31). Indeed, Lynch et al. (27) found a
mg/wound) returned the force absorbed by wounds on combination of PDGF-2 and IGF-I to be the most potent
steroid-treated animals to levels observed in normal inducer for healing partial-thickness porcine wounds
animals and resulted in a histological picture similar to compared with various other individual growth factors
that seen on sections from control wounds on normal or combinations tested.
animals. Although some studies have shown the maximum
The ability of MBWE to promote wound repair was wound strength achieved by the application of single
concentration dependent. Higher concentrations of growth factors (principally TGF-b) to be greater than
MBWE promoted fibroblast influx but did not increase that observed in our experiments, it should be noted
the strength of incisional wounds on either the normal that comparative studies have generally added micro-
or steroid-treated animals. Whereas we cannot rule out gram quantities of single recombinant growth factors to
the presence of some inhibitory activity in MBWE, it is wounds (27, 34, 36, 39, 40). The amount of individual
possible this phenomenon is due to the bimodal action growth factors applied to wounds treated with MBWE
of some individual growth factors in MBWE. Both was in the low nanogram range. It is possible further
TGF-b and bFGF have been shown to have apparent purification of MBWE designed to concentrate specific
concentration-dependent bimodal effects on the strength growth factor components of the extract may enhance
of incisional wounds in vivo (36, 39). Similarly, a the wound healing activity of these fractions.
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR R1659
The current report represents the first to describe the 2. Belford DA, Rogers ML, Francis GL, Payne C, Ballard FJ,
ability of a mitogenic extract containing the comple- and Goddard C. Platelet-derived growth factor, insulin-like
growth factors, fibroblast growth factors and transforming growth
ment of growth factors present in milk to stimulate a factor b do not account for the cell growth activity present in
healing response in organotypic wound models and bovine milk. J Endocrinol 154: 45–55, 1997.
promote healing of the incisional wound in vivo. To- 3. Belford DA, Rogers ML, Regester GO, Francis GL, Smith-
gether with in vitro data showing significant activity in ers GW, Liepe IJ, Priebe IK, and Ballard FJ. Milk-derived
assays of cell growth (3, 10), our results would suggest growth factors as serum supplements for the growth of fibroblast
and epithelial cells. In Vitro Cell Develop Biol 31: 752–760, 1995.
the unique mix of agents in MBWE may confer an 4. Bell E, Ivarsson B, and Merrill C. Production of a tissue-like
ability to promote wound repair over and above that structure by contraction of collagen lattices by human fibroblasts
obtained with single growth factors and that is particu- of different proliferative potential in vitro. Proc Natl Acad Sci
larly relevant to rectifying the healing deficits associ- USA 76: 1274–1278, 1979.
ated with compromised conditions resulting in chronic 5. Bennett NT and Schultz GS. Growth factors and wound
healing: biochemical properties of growth factors and their
cutaneous wounds (47). receptors. Am J Surg 165: 728–737, 1993.
6. Bennett NT and Schultz GS. Growth factors and wound
Perspectives
healing: II. Role in normal and chronic wound healing. Am J
Wound healing is a complex series of events aimed at Surg 166: 74–81, 1993.
7. Brown RL, Breeden MP, and Greenhalgh DG. PDGF and
restoring the integrity of damaged tissue. Included in TGF-a act synergistically to improve wound healing in the
these events are the actions of growth factors, which genetically diabetic mouse. J Surg Res 56: 562–570, 1994.
play an essential role in directing wound repair. The 8. Cook JJ, Haynes KM, and Werther GA. Mitogenic effects of
action of such mediators, however, is disturbed in growth hormone in cultured human fibroblasts; evidence for
action via local insulin-like growth factor I production. J Clin
tissue compromised by lack of blood flow or systemic Invest 81: 206–212, 1988.
diseases such as diabetes, leading to failure of the 9. Cromack DT, Porras-Reyes B, Purdy JA, Pierce GF, and
cellular processes of healing. Whereas growth factors Mustoe TA. Acceleration of tissue repair by transforming growth
have been shown to enhance all aspects of wound repair factor b1: identification of in vivo mechanism of action with
in experimental models, the effectiveness of individual radiotherapy-induced specific healing deficits. Surgery 113: 36–
42, 1993.
growth factors has generally been difficult to replicate 10. Francis GL, Regester GO, Webb HA, and Ballard FJ.
when applied to clinical studies in chronic wounds. As Extraction from cheese whey by cation-exchange chromatogra-
wound repair results from the temporally coordinated phy of factors that stimulate the growth of mammalian cells. J
local expression of multiple families of growth factors Dairy Res 78: 1209–1218, 1995.
and their receptors, it would seem reasonable that the 11. Francis GL, Upton FM, Ballard FJ, McNeil KA, and Wal-
lace JC. Insulin-like growth factors 1 and 2 in bovine colostrum:
application of mixtures of growth factors to the healing sequences and biological activities compared with those of a
wound may be more effective than a single factor. potent truncated form. Biochem J 251: 95–103, 1988.
Bovine milk is a rich source of several classes of growth 12. Fuller GC and Cutroneo KR. Pharmacological interventions.
factors, and cation-exchange fractionation of bovine In: Wound Healing Biochemical and Clinical Aspects, edited by
whey results in a mitogenic extract that contains a 100- Cohen IK, Diegelmann RF, and Lindblad WJ. Philadelphia, PA:
Saunders, 1992, p. 305–315.
to 200-fold enrichment of the growth factor activity 13. Gillery P, Leperre A, Maquart FX, and Borel JP. Insulin-like
present in milk. Our results demonstrate that this growth factor-I (IGF-I) stimulates protein synthesis and collagen
mitogenic extract derived from milk has considerable gene expression in monolayer and lattice cultures of fibroblasts.
wound healing activity, and, given the mixture of J Cell Physiol 152: 389–396, 1992.
growth factors present, we speculate that it may have 14. Gillery P, Maquart FX, and Borel JP. Fibronectin dependence
of the contraction of collagen lattices by human skin fibroblasts.
potential as a clinical treatment to stimulate healing of Exp Cell Res 167: 29–37, 1986.
chronic ulcers and other problem wounds. The socioeco- 15. Grainger DJ, Wakefield L, Bethell HW, Farndale RW, and
nomic benefits of such an agent that can effectively heal Metcalf JC. Release and activation of platelet latent TGF-beta
chronic wounds is likely to be substantial, given the in blood clots during dissolution with plasmin. Nat Med 1:
932–937, 1995.
morbidity and high cost of treatment of this often 16. Greenhalgh DG, Hummel RP III, Albertson S, and Bree-
age-related problem. don MP. Synergistic actions of platelet-derived growth factor
We thank Kaylene Pickering, Xenia Georgiou, Anna Seamark, and the insulin-like growth factors in vivo. Wound Rep Reg 1:
Caroline Payne, and Sue Millard for technical assistance. Microbio- 69–81, 1993.
logical screening of MBWE was undertaken by the Victorian Institute 17. Greenhalgh DG, Sprugel KH, Murray MJ, and Ross R.
of Animal Sciences. PDGF and FGF stimulate wound healing in the genetically
This work was supported by a grant from the Dairy Research and diabetic mouse. Am J Pathol 136: 1235–1246, 1990.
Development Corporation of Australia and the Cooperative Research 18. Grotendorst GR, Martin GR, Pencev D, Sodek J, and
Centre Scheme, Commonwealth Government of Australia. Harvey AK. Stimulation of granulation tissue formation by
Address for reprint requests and other correspondence: T. E. platelet-derived growth factor in normal and diabetic rats. J Clin
Rayner, Child Health Research Institute, Women’s and Children’s Invest 76: 2323–2329, 1985.
Hospital, 72 King William Rd., North Adelaide 5006, SA, Australia 19. Hennessey PJ, Black CT, and Andrassy RJ. Epidermal
(E-mail: trayner@medicine.adelaide.edu.au). growth factor and insulin act synergistically during diabetic
healing. Arch Surg 125: 926–929, 1990.
Received 3 June 1999; accepted in final form 3 December 1999. 20. Ignotz RA and Massagué J. Transforming growth factor-b
stimulates the expression of fibronectin and collagen and their
REFERENCES incorporation into the extracellular matrix. J Biol Chem 261:
1. Belford DA. The mechanism of excisional fetal wound repair in 4337–4345, 1986.
vitro is responsive to growth factors. Endocrinology 138: 3987– 21. Jin Y, Cox DA, Knecht R, Raschdorf F, and Cerletti N.
3996, 1997. Separation, purification, and sequence identification of TGF-beta
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
R1660 MITOGENIC WHEY EXTRACT STIMULATES WOUND REPAIR
1 and TGF-beta 2 from bovine milk. J Protein Chem 10: 565–575, 37. Owens PC, Johnson RJ, Campbell RG, and Ballard FJ.
1991. Growth hormone increases insulin-like growth factor-I (IGF-I)
22. Jyung RW, Mustoe TA, Busby WH, and Clemmons DR. and decreases IGF-II in plasma of growing pigs. J Endocrinol
Increased wound-breaking strength induced by insulin-like 124: 269–275, 1990.
growth factor I in combination with insulin-like growth factor 38. Pierce GF, Mustoe TA, Lingelbach J, Masakowski VR,
binding protein-1. Surgery 115: 233–239, 1994. Gramates P, and Deuel TF. Transforming growth factor b
23. Karr BP, Bubak PJ, Sprugel KH, Pavlin EG, and Engrav reverses the glucocorticoid-induced wound-healing deficit in
LH. Platelet-derived growth factor and wound contraction in the rats: possible regulation in macrophages by platelet-derived
rat. J Surg Res 59: 739–742, 1995. growth factor. Proc Natl Acad Sci USA 86: 2229–2233, 1989.
24. Kucukcelebi A, Carp SS, Hayward PG, Hui P, Cowan WT, 39. Pierce GF, Mustoe TA, Lingelbach J, Masakowski VR,
Ko F, Cooper DM, and Robson MC. Granulocyte-macrophage Griffin GL, Senior RM, and Deuel TF. Platelet-derived growth
colony stimulating factor reverses the inhibition of wound contrac-
factor and transforming growth factor-b enhance tissue repair
tion caused by bacterial contamination. Wounds 4: 241–247,
activities by unique mechanisms. J Cell Biol 109: 429–440, 1989.
1992.
25. Lawrence WT, Sporn MB, Gorschboth C, Norton JA, and 40. Pierce GF, Tarpley JE, Yanagihara D, Mustoe TA, Fox GM,
Grotendorst GR. The reversal of an adriamycin induced heal- and Thomason A. Platelet-derived growth factor (BB ho-
ing impairment with chemoattractants and growth factors. Ann modimer), transforming growth factor-b1, and basic fibroblast
Surg 203: 142–147, 1985. growth factor in dermal wound healing. Am J Pathol 140:
26. LeGrand EK, Senter LH, Gamelli RL, and Kiorpes TC. 1375–1388, 1992.
Evaluation of PDGF-BB, PDGF-AA, bFGF, IL-1, and EGF dose 41. Piscatelli SJ, Michaels BM, Gregory P, Jennings RW,
responses in polyvinyl alcohol sponge implants assessed by a Longaker MT, Harrison MR, and Siebert JW. Fetal fibro-
rapid histologic method. Growth Factors 8: 315–329, 1993. blast contraction of collagen matrices in vitro: the effects of
27. Lynch SE, Colvin RB, and Antoniades HN. Growth factors in epidermal growth factor and transforming growth factor-beta.
wound healing: single and synergistic effects on partial thickness Ann Plast Surg 33: 38–45, 1994.
porcine skin wounds. J Clin Invest 84: 640–646, 1989. 42. Roberts AB, Sporn MB, Assoian RK, Smith JM, Roche NS,
28. Martin P and Lewis J. Actin cables and epidermal movement Wakefield LM, Heine UI, Liotta LA, Falanga V, Kehrl JH,
in embryonic wound healing. Nature 360: 179–183, 1992. and Fauci AS. Transforming growth factor type b: rapid induc-
29. Mast BA and Schultz GS. Interactions of cytokines, growth tion of fibrosis and angiogenesis in vivo and stimulation of
factors and proteases in acute and chronic wounds. Wound Rep collagen formation in vitro. Proc Natl Acad Sci USA 83: 4167–
Reg 4: 411–420, 1996. 4171, 1986.
30. McGee GS, Davidson JM, Buckley A, Sommer A, Wood- 43. Robson MC. The role of growth factors in the healing of chronic
ward SC, Aquino AM, Barbour R, and Demetriou AA. wounds. Wound Rep Reg 5: 12–17, 1997.
Recombinant basic fibroblast growth factor accelerates wound 44. Rogers ML, Belford DA, Francis GL, and Ballard FJ.
healing. J Surg Res 45: 145–153, 1988. Identification of fibroblast growth factors in bovine cheese whey.
31. Mellin TN, Mennie RJ, Cashen DE, Ronan JJ, Capparella J Dairy Res 62: 501–507, 1995.
J, James ML, DiSalvo J, Frank J, Linemeyer D, Gimenez- 45. Rogers ML, Goddard C, Regester GO, Ballard FJ, and
Gallego G, and Thomas KA. Acidic fibroblast growth factor Belford DA. Transforming growth factor b in bovine milk:
accelerates dermal wound healing. Growth Factors 7: 1–14, concentration, stability and molecular mass forms. J Endocrinol
1992. 151: 77–86, 1996.
32. Montesano R and Orci L. Transforming growth factor beta 46. Shing Y and Klagsbrun M. Purification and characterization of
stimulates collagen-matrix contraction by fibroblasts: implica-
a bovine colostrum-derived growth factor. Mol Endocrinol 1:
tions for wound healing. Proc Natl Acad Sci USA 85: 4894–4897,
335–338, 1987.
1988.
47. Stadelmann WK, Digenis AG, and Tobin GR. Physiology and
33. Mueller RV, Spencer EM, Sommer A, Maack CA, Suh D,
and Hunt TK. The role of IGF-I and IGFBP-3 in wound healing. healing dynamics of chronic cutaneous wounds. Am J Surg 176,
In: Modern Concepts of Insulin-like Growth Factors, edited by Suppl 2A: 26S-38S, 1998.
Spencer EM. New York: Elsevier, 1991, p. 185–192. 48. Suh DY, Hunt TK, and Spencer EM. Insulin-like growth
34. Mustoe TA, Pierce GF, Thomason A, Gramates P, Sporn factor-I reverses the impairment of wound healing induced by
MB, and Deuel TF. Accelerated healing of incisional wounds in corticosteroids in rats. Endocrinology 131: 2399–2403, 1992.
rats induced by transforming growth factor-b. Science 237: 49. Uhl E, Barker JH, Bondàr I, Galla TJ, Leiderer R, Lehr HA,
1333–1336, 1987. and Messmer K. Basic fibroblast growth factor accelerates
35. Mustoe TA, Purdy J, Gramates P, Deuel TF, Thomason A, wound healing in chronically ischaemic tissue. Br J Surg 80:
and Pierce GF. Reversal of impaired wound healing in irradi- 977–980, 1993.
ated rats by platelet-derived growth factor-BB. Am J Surg 158: 50. Yager DR, Chen SM, Ward SI, Olutoye OO, Diegelmann RF,
345–350, 1989. and Cohen IK. Ability of chronic wound fluids to degrade
36. Okumura M, Okuda T, Nakamura T, and Yajima M. Effect of peptide growth factors is associated with increased levels of
basic fibroblast growth factor on wound healing in healing- elastase activity and diminished levels of proteinase inhibitors.
impaired animal models. Drug Res 46: 547–551, 1996. Wound Rep Reg 5: 23–32, 1997.
Downloaded from journals.physiology.org/journal/ajpregu (114.125.248.089) on December 13, 2020.
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Pathophysiology Test Bank CH 6-9Document33 pagesPathophysiology Test Bank CH 6-9Joyy86% (7)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Krok Book PDFDocument1,282 pagesKrok Book PDFAnasse Douma100% (1)
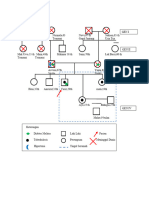
- GenogramDocument1 pageGenogramHaykal Estu BhismoroNo ratings yet
- 377-Article Text-1099-1-10-20191009Document16 pages377-Article Text-1099-1-10-20191009Haykal Estu BhismoroNo ratings yet
- Mucus ServixDocument10 pagesMucus ServixHaykal Estu BhismoroNo ratings yet
- Transverse Myelitis - Johns Hopkins MedicineDocument6 pagesTransverse Myelitis - Johns Hopkins MedicineHaykal Estu BhismoroNo ratings yet
- FIGO (International Federation Obstetrics and Gynecology) Pre-Eclampsia 2019Document58 pagesFIGO (International Federation Obstetrics and Gynecology) Pre-Eclampsia 2019Haykal Estu BhismoroNo ratings yet
- HHS Public Access: Wound Repair and Regeneration: Mechanisms, Signaling, and TranslationDocument36 pagesHHS Public Access: Wound Repair and Regeneration: Mechanisms, Signaling, and TranslationHaykal Estu BhismoroNo ratings yet
- H2 Cell Structure QuestionsDocument15 pagesH2 Cell Structure QuestionsWesley Tan100% (2)
- Muscular TissueDocument49 pagesMuscular TissueSteph VeeNo ratings yet
- Addis Ababa University Institute of Biotechnology Presentation Assignment For The Course Advanced Molecular Biology (BIOT 801)Document23 pagesAddis Ababa University Institute of Biotechnology Presentation Assignment For The Course Advanced Molecular Biology (BIOT 801)HelenNo ratings yet
- Chemistry of FragranceDocument6 pagesChemistry of FragrancePélagie TuyishimireNo ratings yet
- Topic 6 Gene Control: Suggested ActivitiesDocument2 pagesTopic 6 Gene Control: Suggested ActivitiesSemwezi EnockNo ratings yet
- Chapter 4Document29 pagesChapter 4skywalkerNo ratings yet
- GR 6-CELL - LessonDocument27 pagesGR 6-CELL - LessonshamshadNo ratings yet
- Bacterial Antibiotics: Mechanisms of ResistanceDocument10 pagesBacterial Antibiotics: Mechanisms of ResistanceAnnisa YohanesNo ratings yet
- Marco TosDocument1 pageMarco TosMarco Bernabe JumaquioNo ratings yet
- Cloning and Expression - FLAG and 3xFLAG OverviewDocument3 pagesCloning and Expression - FLAG and 3xFLAG OverviewSigma-Aldrich100% (12)
- Wardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test BankDocument38 pagesWardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test Bankedricduyenuc1uw100% (25)
- Mmoi BC SbaDocument15 pagesMmoi BC SbaYoon YoonNo ratings yet
- Q.P. Code: 684884Document1 pageQ.P. Code: 684884Priyanjali SainiNo ratings yet
- Aerobic Respiration Quiz SCDocument2 pagesAerobic Respiration Quiz SCapi-323720899No ratings yet
- A Reappraisal of CTLA-4 Checkpoint Blockade in Cancer ImmunotherapyDocument17 pagesA Reappraisal of CTLA-4 Checkpoint Blockade in Cancer ImmunotherapyKRUBAKARAN MUTHUSAMYNo ratings yet
- ActivityReport IMM2004Document211 pagesActivityReport IMM2004mehhh123No ratings yet
- Soalan Ulangkaji Bab 5 Tingkatan 4 PDFDocument22 pagesSoalan Ulangkaji Bab 5 Tingkatan 4 PDFIsmaliza Ishak0% (1)
- Photosynthesis in Higher PlantsDocument26 pagesPhotosynthesis in Higher PlantsShadab HanafiNo ratings yet
- Anabolism: The Use of Energy in Biosynthesis: Chapter OverviewDocument5 pagesAnabolism: The Use of Energy in Biosynthesis: Chapter OverviewElishae SamonteNo ratings yet
- Stem Cells TherapyDocument21 pagesStem Cells TherapyHONGJYNo ratings yet
- Vacuole FunctionDocument8 pagesVacuole Functionkbansal981No ratings yet
- CHE631-Module 4 - EnzymesDocument26 pagesCHE631-Module 4 - EnzymesrutwickNo ratings yet
- WEEK3 ModuleLec (BIO101)Document10 pagesWEEK3 ModuleLec (BIO101)Maricris GatdulaNo ratings yet
- The Pathogenic Basis of Malaria: InsightDocument7 pagesThe Pathogenic Basis of Malaria: InsightRaena SepryanaNo ratings yet
- Chymosin ReninDocument1 pageChymosin ReninArfanizam Bin ArisNo ratings yet
- PhysiologyWorkbook7 5Document267 pagesPhysiologyWorkbook7 5SophiaNo ratings yet
- Origami Protein HandoutDocument4 pagesOrigami Protein HandoutRaajeswaran BaskaranNo ratings yet
- Porphyromonas Gingivalis: An: Overview of Periodontopathic Pathogen Below The Gum LineDocument14 pagesPorphyromonas Gingivalis: An: Overview of Periodontopathic Pathogen Below The Gum LineSasa AprilaNo ratings yet