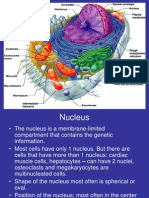
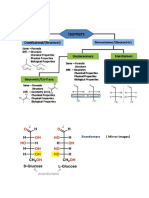
You might also like
- Types of TransposonsDocument13 pagesTypes of Transposonsmozhi74826207No ratings yet
- TranscriptionDocument70 pagesTranscriptionkhan aishaNo ratings yet
- Biotecnology and Its ApplicationDocument87 pagesBiotecnology and Its ApplicationRangaswamyBiligiraiah100% (1)
- Linkage: Harshraj Subhash Shinde KKW, Cabt, NashikDocument14 pagesLinkage: Harshraj Subhash Shinde KKW, Cabt, Nashiksivaram888No ratings yet
- Replication in EukaryotesDocument8 pagesReplication in EukaryotesnaliniNo ratings yet
- RFLP & RapdDocument25 pagesRFLP & RapdBalaji Prasanna KumarNo ratings yet
- DNA Manipulative EnzymesDocument17 pagesDNA Manipulative EnzymesZain Ul AbedienNo ratings yet
- cDNA Libraries and Gene CloningDocument8 pagescDNA Libraries and Gene CloningRoberto RomeroNo ratings yet
- Genome Organization 1Document42 pagesGenome Organization 1nsjunnarkar100% (1)
- Gene Silencing: An In-Depth Look at Mechanisms and ApplicationsDocument18 pagesGene Silencing: An In-Depth Look at Mechanisms and ApplicationsNAMRA RASHEEDNo ratings yet
- Transcription and RNA ProcessingDocument38 pagesTranscription and RNA ProcessingRishi Kumar100% (1)
- Agrobacterium Mediated Gene Transfer BA3825 SWaghmareDocument8 pagesAgrobacterium Mediated Gene Transfer BA3825 SWaghmareSubash Ragasudha100% (1)
- 5.1 Blotting TechniquesDocument33 pages5.1 Blotting TechniquesAmir Al-AimanNo ratings yet
- Transcription in ProkaryotesDocument16 pagesTranscription in ProkaryotesAditya Kanwal100% (1)
- Post Translational ModificationDocument4 pagesPost Translational ModificationSaira DogarNo ratings yet
- What is blotting techniquesDocument28 pagesWhat is blotting techniquesShivraj JadhavNo ratings yet
- Genomes in Prokaryotes and EukaryotesDocument46 pagesGenomes in Prokaryotes and EukaryoteskityamuwesiNo ratings yet
- Post Transcriptional ModificationDocument31 pagesPost Transcriptional ModificationranasiddharthNo ratings yet
- M.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarDocument39 pagesM.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarM.PRASAD NAIDUNo ratings yet
- Lecture 1 - Chromosome & GenomeDocument64 pagesLecture 1 - Chromosome & GenomeFriendlyGoodGirlNo ratings yet
- Gene Silencing: Presented by Aastha Pal M.Sc. 4 Semester (Biotechnology) Swami Rama Himalayan UniversityDocument22 pagesGene Silencing: Presented by Aastha Pal M.Sc. 4 Semester (Biotechnology) Swami Rama Himalayan UniversityAmit NegiNo ratings yet
- Cell Cycle and Cell DivisionDocument12 pagesCell Cycle and Cell DivisionMajid Al-hachami100% (1)
- Regulation of Gene Expression in Prokaryotes: © John Wiley & Sons, IncDocument65 pagesRegulation of Gene Expression in Prokaryotes: © John Wiley & Sons, IncRoberto CastroNo ratings yet
- Central DogmaDocument35 pagesCentral Dogmatariqul13017No ratings yet
- Inheritance Biology EDUNCLEDocument54 pagesInheritance Biology EDUNCLEEvita Almeida100% (1)
- Postranslational ModificationDocument78 pagesPostranslational ModificationnsjunnarkarNo ratings yet
- Restriction EnzymeDocument33 pagesRestriction EnzymeFawaz MohamadNo ratings yet
- Submitted by Aiswarya V 1St MSC Zoology Roll Number 3301Document32 pagesSubmitted by Aiswarya V 1St MSC Zoology Roll Number 3301PES PEOPLENo ratings yet
- 09 Regulation of Gene ExpressionDocument42 pages09 Regulation of Gene ExpressionFirst Anonym877No ratings yet
- The Nucleus and Nuclear EnvelopeDocument58 pagesThe Nucleus and Nuclear EnvelopeImperiall100% (1)
- Unit 7 Operon ConceptDocument18 pagesUnit 7 Operon ConceptSarah PavuNo ratings yet
- Bacterial, GeneticsDocument21 pagesBacterial, GeneticsMoona WahabNo ratings yet
- MOL 125 Lecture 5&6 RNA Transcription: Lippincott's Illustrated Reviews: Cell and Molecular Biology, DR Hanaa HajeerDocument52 pagesMOL 125 Lecture 5&6 RNA Transcription: Lippincott's Illustrated Reviews: Cell and Molecular Biology, DR Hanaa HajeerAffan Elahi100% (1)
- VectorsDocument6 pagesVectorsAssad MustafaNo ratings yet
- General Translation MechanismDocument15 pagesGeneral Translation MechanismAishwarya KashyapNo ratings yet
- DNA Replication in Prokaryotes and EukaryotesDocument7 pagesDNA Replication in Prokaryotes and Eukaryotessmn416No ratings yet
- Enzymes and Proteins in Dna ReplicationDocument39 pagesEnzymes and Proteins in Dna ReplicationRININo ratings yet
- Staining Techniques in Biological StudiesDocument52 pagesStaining Techniques in Biological StudiesRamyaAbhishekShenoyNo ratings yet
- Dna Sequencing: DR Z Chikwambi BiotechnologyDocument110 pagesDna Sequencing: DR Z Chikwambi BiotechnologyMutsawashe Munetsi100% (1)
- Post-Transcriptional Modification: An Overview of mRNA ProcessingDocument20 pagesPost-Transcriptional Modification: An Overview of mRNA ProcessingZain YaqoobNo ratings yet
- Gene Transfer TechniquesDocument36 pagesGene Transfer TechniquesDrMumtaz F MusaliarNo ratings yet
- Transcription and TranslationDocument9 pagesTranscription and TranslationlinhNo ratings yet
- microRNA, Gene Mapping and Gene SequencingDocument33 pagesmicroRNA, Gene Mapping and Gene SequencingAdarsh ShrivastavaNo ratings yet
- DNA Organization in Eukaryotic Chromosomes: Chapter 12: Section 12.4Document22 pagesDNA Organization in Eukaryotic Chromosomes: Chapter 12: Section 12.4Jaisy Samuel100% (1)
- Dna SupercoilingDocument16 pagesDna SupercoilingrainabtNo ratings yet
- Prokaryotic Genome OrganizationDocument53 pagesProkaryotic Genome OrganizationAqsa YaminNo ratings yet
- Regulation of Gene ExpressionDocument18 pagesRegulation of Gene Expressionchocoholic potchiNo ratings yet
- Construction of Genomic and cDNA Libraries-AmitDocument10 pagesConstruction of Genomic and cDNA Libraries-AmitPrashant BajpaiNo ratings yet
- Blotting TechniqueDocument34 pagesBlotting TechniqueUmair100% (1)
- Process of Transcription in Eukaryotes ExplainedDocument1 pageProcess of Transcription in Eukaryotes ExplainedSomNo ratings yet
- Genetic Code HMDocument30 pagesGenetic Code HMdrhmpatel100% (2)
- DNA Replication: How DNA Is CopiedDocument13 pagesDNA Replication: How DNA Is CopieddvdmegaNo ratings yet
- CentrifugationDocument20 pagesCentrifugationMuhammad Asif ShaheenNo ratings yet
- Host Parasite RelationshipDocument12 pagesHost Parasite RelationshipAyesha MasoodNo ratings yet
- Day 3 - Transcription and RNA ProcessingDocument50 pagesDay 3 - Transcription and RNA ProcessingAniket ParabNo ratings yet
- Bacterial ConjugationDocument15 pagesBacterial ConjugationNiveditha ChalukyaNo ratings yet
- Composed of A BaseDocument23 pagesComposed of A Basetanvi bhallaNo ratings yet
- In-Depth Steps Towards Nucleic Acid and Protein SynthesisDocument21 pagesIn-Depth Steps Towards Nucleic Acid and Protein SynthesisGbenga AjaniNo ratings yet
- Transgenic AnimalsDocument13 pagesTransgenic AnimalsSamunderSinghSinghraNo ratings yet
- Carbohydrates and Their Role in Future Time ManagementDocument3 pagesCarbohydrates and Their Role in Future Time ManagementBandook GamersNo ratings yet
- Enantiomers: (Mirror Images)Document5 pagesEnantiomers: (Mirror Images)Bandook GamersNo ratings yet
- 6-Regulation of Gene Expression in ProkaryotesDocument22 pages6-Regulation of Gene Expression in ProkaryotesJasdeep KaurNo ratings yet
- CarbohydratesDocument4 pagesCarbohydratesBandook GamersNo ratings yet
- Concept of Gene PoolDocument41 pagesConcept of Gene PoolBandook Gamers0% (1)
- Recap of key microbiology conceptsDocument39 pagesRecap of key microbiology conceptsMehrin KabirNo ratings yet
- BASIC VIROLOGY, Third Edition by Wagner, Hewlett, Bloom and CameriniDocument25 pagesBASIC VIROLOGY, Third Edition by Wagner, Hewlett, Bloom and CameriniJason PhalangeNo ratings yet
- SEMINAR-Clinical Microbiology: Topic-Human Immuno-Deficiency Virus (HIV)Document54 pagesSEMINAR-Clinical Microbiology: Topic-Human Immuno-Deficiency Virus (HIV)sushant_dharNo ratings yet
- ISO 20395:2019(en) - qPCR and dPCR quantification methodsDocument44 pagesISO 20395:2019(en) - qPCR and dPCR quantification methodsGilberto Ragnar Nodarse100% (1)
- HIV & AIDS PathophysiologyDocument3 pagesHIV & AIDS PathophysiologyMarinill Soliman100% (3)
- Acellular Life and VirusesDocument4 pagesAcellular Life and VirusesRahila AkramNo ratings yet
- Biodiversity, Classification and ConservationDocument17 pagesBiodiversity, Classification and Conservationmobasser sikderNo ratings yet
- CH17Document86 pagesCH17y9bt429No ratings yet
- Genetic EngineeringDocument27 pagesGenetic EngineeringSeema QureshiNo ratings yet
- Virology SyllabusDocument6 pagesVirology SyllabusRichard S. RoxasNo ratings yet
- Diagnosing COVID-19: The Disease and Tools For Detection: AccessDocument14 pagesDiagnosing COVID-19: The Disease and Tools For Detection: AccessYurley TatianaNo ratings yet
- Heteroduplex Formation and S1 Digestion For Mapping Alternative Splicing SitesDocument12 pagesHeteroduplex Formation and S1 Digestion For Mapping Alternative Splicing SitesCleaver BrightNo ratings yet
- Module 6 RationaleDocument1 pageModule 6 RationaleG INo ratings yet
- GSE171524 Series MatrixDocument17 pagesGSE171524 Series MatrixCong Trang MaiNo ratings yet
- Procleix Ultrio Plus AssayDocument61 pagesProcleix Ultrio Plus AssaydatitoxNo ratings yet
- Directorate of Learning Systems Directorate of Learning SystemsDocument50 pagesDirectorate of Learning Systems Directorate of Learning SystemsMatin Ahmad KhanNo ratings yet
- Soluzioni Esercizi Del LibroDocument8 pagesSoluzioni Esercizi Del Librofeysalnur100% (1)
- HIV - PathogenesisDocument14 pagesHIV - Pathogenesisjunaidiabdhalim100% (1)
- Gene Expression 1Document14 pagesGene Expression 1John LazaroNo ratings yet
- PCR Technology Current Innovations Third EditionDocument453 pagesPCR Technology Current Innovations Third Editiontkn5520No ratings yet
- DNA Polymerase IIIDocument24 pagesDNA Polymerase IIIAhmad JavaidNo ratings yet
- Understanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionDocument27 pagesUnderstanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionNikhil BijuNo ratings yet
- Test Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadDocument23 pagesTest Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadMeganGomezpizb100% (17)
- Description: Sensifast™ Cdna Synthesis KitDocument2 pagesDescription: Sensifast™ Cdna Synthesis KitDevin HendrawanNo ratings yet
- Genetic Terminology ExplainedDocument6 pagesGenetic Terminology ExplainedMuthu KaruppanNo ratings yet
- Sprin 1 PDFDocument177 pagesSprin 1 PDFSoumashree MondalNo ratings yet
- Catálogo PromegaDocument317 pagesCatálogo PromegaMon Era100% (1)
- Genetic Engineering and Recombinant DNA TechnologyDocument38 pagesGenetic Engineering and Recombinant DNA Technologyseada JemalNo ratings yet
- EIA Chapter PDFDocument29 pagesEIA Chapter PDFSantiago ValeroNo ratings yet
- GAT B 2021 Practice QuestionsDocument29 pagesGAT B 2021 Practice QuestionsVaishnavi ReddyNo ratings yet