You might also like
- Clinical Manual of Blood and Bone Marrow TransplantationFrom EverandClinical Manual of Blood and Bone Marrow TransplantationSyed A. AbutalibNo ratings yet
- Coagulation PathwayDocument10 pagesCoagulation Pathwayamonra10No ratings yet
- Platelet Function - Assessment, Diagnosis, and TreatmentDocument400 pagesPlatelet Function - Assessment, Diagnosis, and TreatmentethainkeroNo ratings yet
- Hemostasis and Thrombosis: Practical Guidelines in Clinical ManagementFrom EverandHemostasis and Thrombosis: Practical Guidelines in Clinical ManagementHussain I. SabaNo ratings yet
- Platelet Function Analysis PDFDocument13 pagesPlatelet Function Analysis PDFDiah Puspita RiniNo ratings yet
- Biomechanics and Related Bio-Engineering Topics: Proceedings of a Symposium Held in Glasgow, September 1964From EverandBiomechanics and Related Bio-Engineering Topics: Proceedings of a Symposium Held in Glasgow, September 1964No ratings yet
- Antibodies Safety PDFDocument14 pagesAntibodies Safety PDFTai BuiNo ratings yet
- Cancer Cytogenetics: Chromosomal and Molecular Genetic Aberrations of Tumor CellsFrom EverandCancer Cytogenetics: Chromosomal and Molecular Genetic Aberrations of Tumor CellsNo ratings yet
- PS40 BloodMarrow Booklet 2023Document72 pagesPS40 BloodMarrow Booklet 2023wilkesgillinghamNo ratings yet
- Tellurium in Organic Synthesis: Second, Updated and Enlarged EditionFrom EverandTellurium in Organic Synthesis: Second, Updated and Enlarged EditionNo ratings yet
- CytotoxicityDocument15 pagesCytotoxicityLama El NajdiNo ratings yet
- Hemostasis and Transfusion Medicine GuideDocument83 pagesHemostasis and Transfusion Medicine GuideJesserene Mangulad SorianoNo ratings yet
- Sca PDFDocument58 pagesSca PDFMUHAMMAD09No ratings yet
- How I Use Platelet TransfusionsDocument12 pagesHow I Use Platelet TransfusionsJovanna OrtuñoNo ratings yet
- Sarthak Mechanism of Blood ClottingDocument42 pagesSarthak Mechanism of Blood ClottingVishal MishraNo ratings yet
- Immuno2 Cells of The Immune SystemDocument36 pagesImmuno2 Cells of The Immune Systemmmoney1No ratings yet
- Dual Antiplatelet Therapy for Coronary and Peripheral Arterial DiseaseFrom EverandDual Antiplatelet Therapy for Coronary and Peripheral Arterial DiseaseSorin BrenerNo ratings yet
- Michigan Nurses Association v. Huron Valley-Sinai HospitalDocument23 pagesMichigan Nurses Association v. Huron Valley-Sinai HospitalFindLawNo ratings yet
- Transfusion: Practices in NicuDocument80 pagesTransfusion: Practices in NicuPranay MallNo ratings yet
- Dental Problems in Primary CareDocument7 pagesDental Problems in Primary Carehawraa tarhiniNo ratings yet
- Multiple MyelomaDocument287 pagesMultiple MyelomaMarco Antonio Grajales BuitragoNo ratings yet
- Review of Current Transfusion Therapy and Blood Banking PracticesDocument52 pagesReview of Current Transfusion Therapy and Blood Banking PracticesAmaranto SantosoNo ratings yet
- Leukemia and Lymphoma Imaging Advances in Pediatric Cancer ManagementDocument31 pagesLeukemia and Lymphoma Imaging Advances in Pediatric Cancer ManagementPartha GanesanNo ratings yet
- FFP and Cryoprecipitate FinalDocument6 pagesFFP and Cryoprecipitate FinalganotNo ratings yet
- Transfusión IntraoperatoriaDocument24 pagesTransfusión IntraoperatoriaGiovany Salinas100% (1)
- Short and Long Term Regulation and Determinants of BPDocument90 pagesShort and Long Term Regulation and Determinants of BPDan Tristam MicabaloNo ratings yet
- History Anesthesia 2003-2007Document9 pagesHistory Anesthesia 2003-2007John RaufNo ratings yet
- Essentials of Internal MedicineDocument832 pagesEssentials of Internal MedicineHtet Kyaw KyawNo ratings yet
- Lacks v. ThermoFisherDocument15 pagesLacks v. ThermoFisherjssherkowNo ratings yet
- Cellular Injury, Adaptation and Cellular DeathDocument40 pagesCellular Injury, Adaptation and Cellular DeathSuleiman KikulweNo ratings yet
- Immnunology Notebook Chapter One: Innate ImmunityDocument45 pagesImmnunology Notebook Chapter One: Innate ImmunityJavier Alejandro Daza GalvánNo ratings yet
- Approach To Refractory Childhood SeizuresDocument10 pagesApproach To Refractory Childhood SeizureskholisahnasutionNo ratings yet
- Best Physiology TextbookDocument3 pagesBest Physiology TextbookPeterNo ratings yet
- Aabb PBM WhitepaperDocument18 pagesAabb PBM WhitepaperdamadolNo ratings yet
- 2018 Highlights of ASH in Asia Pacific Program BookDocument415 pages2018 Highlights of ASH in Asia Pacific Program BookSyavira EzizaNo ratings yet
- Antigen AntibodiesDocument65 pagesAntigen AntibodiespradeepbawaneNo ratings yet
- Eiichi Ishii (Eds.) - Hematological Disorders in Children - Pathogenesis and Treatment-Springer Singapore (2017)Document268 pagesEiichi Ishii (Eds.) - Hematological Disorders in Children - Pathogenesis and Treatment-Springer Singapore (2017)ZoraRosaNo ratings yet
- Gastrointestinal DiseasesDocument853 pagesGastrointestinal DiseasesCiobanasu Denisa100% (1)
- Immunophenotyping For HematologistDocument139 pagesImmunophenotyping For HematologistMantavya PurohitNo ratings yet
- Opportunities and Challenges in Biosimilar Development: Focus N..Document8 pagesOpportunities and Challenges in Biosimilar Development: Focus N..mgarbini8361No ratings yet
- Clinical Genetics and Genomics Oxford University b8h DR NotesDocument935 pagesClinical Genetics and Genomics Oxford University b8h DR NotesZay Yar WinNo ratings yet
- Eight Hallmarks of Cancer ExplainedDocument40 pagesEight Hallmarks of Cancer ExplainedArnab KalitaNo ratings yet
- Acute and Preventive Pharmacologic Treatment of Cluster HeadacheDocument12 pagesAcute and Preventive Pharmacologic Treatment of Cluster Headachelusi jelitaNo ratings yet
- PlateletsDocument35 pagesPlateletsSiĦãm Mỡ100% (1)
- White Blood CellDocument8 pagesWhite Blood Cellsai calderNo ratings yet
- 1 Juliane Flores Bible FinalDocument170 pages1 Juliane Flores Bible FinalJuliane FloresNo ratings yet
- Haematologists Toolkit V1b August 16 EoEDocument23 pagesHaematologists Toolkit V1b August 16 EoELlrss AdnNo ratings yet
- Cell-Mediated Immune Responses8Document26 pagesCell-Mediated Immune Responses8Trifu AndreiNo ratings yet
- Medication Deferral List ExplainedDocument2 pagesMedication Deferral List ExplainedKwiila KaoNo ratings yet
- Blood Transfusion Therapy DefinitionDocument27 pagesBlood Transfusion Therapy DefinitionEvangelin MelvinNo ratings yet
- Receptor Tyrosine Kinase InhibitorsDocument63 pagesReceptor Tyrosine Kinase InhibitorsNoor Nour100% (1)
- Blood Test RangesDocument18 pagesBlood Test Rangeskltran80No ratings yet
- Hepatitis C SlidesDocument51 pagesHepatitis C SlidesChukwu NedumNo ratings yet
- Blood Safety and SURVEILLANCE PDFDocument629 pagesBlood Safety and SURVEILLANCE PDFsahrish100% (1)
- Southern African Journal of Anaesthesia and AnalgesiaDocument7 pagesSouthern African Journal of Anaesthesia and AnalgesiaRochnald PigaiNo ratings yet
- 1 s2.0 S1319562X19300555 MainDocument4 pages1 s2.0 S1319562X19300555 MainRochnald PigaiNo ratings yet
- Amendt PracticeDocument16 pagesAmendt PracticeRochnald PigaiNo ratings yet
- Investigation of Nocturnal Oviposition byDocument2 pagesInvestigation of Nocturnal Oviposition byRochnald PigaiNo ratings yet
- Complications of Using Laryngeal Mask Airway DurinDocument6 pagesComplications of Using Laryngeal Mask Airway DurinRochnald PigaiNo ratings yet
- Comparison of The Soft Seal Laryngeal Mask in Spontaneously Breathing Adult PatientsDocument6 pagesComparison of The Soft Seal Laryngeal Mask in Spontaneously Breathing Adult PatientsRochnald PigaiNo ratings yet
- 244 / OphthalmologyDocument2 pages244 / OphthalmologyRochnald PigaiNo ratings yet
- Malignant Melanoma of The Eyelid SkinDocument7 pagesMalignant Melanoma of The Eyelid SkinRochnald PigaiNo ratings yet
- Premedication With Fentanyl For Less Invasive SurfDocument19 pagesPremedication With Fentanyl For Less Invasive SurfRochnald PigaiNo ratings yet
- Epidemiological Trends in Malignant Lacrimal Gland TumorsDocument5 pagesEpidemiological Trends in Malignant Lacrimal Gland TumorsRochnald PigaiNo ratings yet
- Noninvasive Identification of Benign and Malignant Eyelid Tumors Using Clinical Images Via Deep Learning SystemDocument14 pagesNoninvasive Identification of Benign and Malignant Eyelid Tumors Using Clinical Images Via Deep Learning SystemRochnald PigaiNo ratings yet
- 01 Sa 0000132004 32857 BCDocument2 pages01 Sa 0000132004 32857 BCRochnald PigaiNo ratings yet
- Masses of The Lacrimal Gland Evaluation and TreatmmentDocument7 pagesMasses of The Lacrimal Gland Evaluation and TreatmmentRochnald PigaiNo ratings yet
- Cornea2006LacrimalGland AnatomyhistopathologyDocument9 pagesCornea2006LacrimalGland AnatomyhistopathologyRochnald PigaiNo ratings yet
- Bmri2015 453091Document5 pagesBmri2015 453091Rochnald PigaiNo ratings yet
- Malignant Melanoma of The Eyelid Skin HistopatholoDocument8 pagesMalignant Melanoma of The Eyelid Skin HistopatholoRochnald PigaiNo ratings yet
- Temperature Sensitivity of Soil Carbon Decomposition and Feedbacks To Climate ChangeDocument9 pagesTemperature Sensitivity of Soil Carbon Decomposition and Feedbacks To Climate ChangeRochnald PigaiNo ratings yet
- Clinical Characteristics and Outcome of Malignant Eyelid Tumors: A Five-Year Retrospective StudyDocument4 pagesClinical Characteristics and Outcome of Malignant Eyelid Tumors: A Five-Year Retrospective StudyRochnald PigaiNo ratings yet
- Surgical Management of Lacrimal Gland TumorsDocument6 pagesSurgical Management of Lacrimal Gland TumorsRochnald PigaiNo ratings yet
- Hemangiomas An OverviewDocument18 pagesHemangiomas An OverviewRochnald PigaiNo ratings yet
- Gunduz - 2018 - Overview of Benign and Malignant Lacrimal Gland TumorsDocument11 pagesGunduz - 2018 - Overview of Benign and Malignant Lacrimal Gland TumorscutfzNo ratings yet
- Vascularaccess - 508 HemoDocument8 pagesVascularaccess - 508 HemoAnnisa KartikasariNo ratings yet
- Entomology and The Law-Flies As Forensic IndicatorsDocument2 pagesEntomology and The Law-Flies As Forensic IndicatorsRochnald PigaiNo ratings yet
- Opr 01 YXUDocument5 pagesOpr 01 YXUAnonymous rQchwzLXANo ratings yet
- The Use of A Cycloheximide-Chloramphenicol MediumDocument5 pagesThe Use of A Cycloheximide-Chloramphenicol MediumRochnald PigaiNo ratings yet
- Glasjpd PPDDocument14 pagesGlasjpd PPDJr Ol FDNo ratings yet
- Giant Hanging Melanoma of The Eye SkinDocument2 pagesGiant Hanging Melanoma of The Eye SkinRochnald PigaiNo ratings yet
- WHO - Haemoglobin Level For Anemia 2011Document6 pagesWHO - Haemoglobin Level For Anemia 2011Adhi SanjayaNo ratings yet
- Antifungal Drugs During Pregnancy An Updated ReviewDocument9 pagesAntifungal Drugs During Pregnancy An Updated ReviewRochnald PigaiNo ratings yet
- Ijcc 2 019 PDFDocument7 pagesIjcc 2 019 PDFAllam SandeepNo ratings yet
- Immunity: by Ashok K TiwariDocument42 pagesImmunity: by Ashok K TiwariXEDGER09No ratings yet
- Learner Guide: Cambridge O Level Biology 5090Document45 pagesLearner Guide: Cambridge O Level Biology 5090Dawood100% (1)
- PMLS 2 PrelimsDocument55 pagesPMLS 2 PrelimsCassey Stephane PinedaNo ratings yet
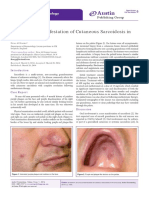
- Austin: An Unusual Manifestation of Cutaneous Sarcoidosis in The Oral CavityDocument2 pagesAustin: An Unusual Manifestation of Cutaneous Sarcoidosis in The Oral CavityRENITA USWATUN HASANAH 1No ratings yet
- Immunology and Serology Lecture # 2: Overview of ImmunityDocument14 pagesImmunology and Serology Lecture # 2: Overview of ImmunityBryan Michael BalunesNo ratings yet
- ZL 124 Lecture 7 - Gastrulation in Birds & MammalsDocument36 pagesZL 124 Lecture 7 - Gastrulation in Birds & MammalsNestory MartineNo ratings yet
- Human Body 11 Organ Systems FoldableDocument4 pagesHuman Body 11 Organ Systems Foldablenaeleti1899No ratings yet
- (TANUVAS) General Veterinary PathologyDocument126 pages(TANUVAS) General Veterinary PathologyKishore SenthilkumarNo ratings yet
- Histo Lab Exercise #14Document4 pagesHisto Lab Exercise #14Gela ReyesNo ratings yet
- Maternal and Child Nursing - Antepartum Practice Test With AnswersDocument6 pagesMaternal and Child Nursing - Antepartum Practice Test With Answersgelean payodNo ratings yet
- Urban - Effectiveness of Two Different Lingual Flap Advancing Techniques For Vertical Bone Augmentation in The Posterior MandibleDocument7 pagesUrban - Effectiveness of Two Different Lingual Flap Advancing Techniques For Vertical Bone Augmentation in The Posterior MandibleEliza DraganNo ratings yet
- G10 Science Q3 The Human Reproductive System - PresentationDocument37 pagesG10 Science Q3 The Human Reproductive System - Presentationma. eleanorNo ratings yet
- Morphology Internationaljournalplantsciences2011Document14 pagesMorphology Internationaljournalplantsciences2011wenagoNo ratings yet
- Skills Input - Leopold's ManeuverDocument13 pagesSkills Input - Leopold's ManeuverJerald FernandezNo ratings yet
- Histology - The Microscopic Anatomy and Function of Tissues and OrgansDocument423 pagesHistology - The Microscopic Anatomy and Function of Tissues and Organsdorina0101100% (1)
- 1 Group Case Study - NCM 107 - Group 3Document9 pages1 Group Case Study - NCM 107 - Group 3Princess Pilove GawongnaNo ratings yet
- PASIEN DIGESTIF OHO - 2 Januari 2019 (R1, R2B, R3, R4B)Document2 pagesPASIEN DIGESTIF OHO - 2 Januari 2019 (R1, R2B, R3, R4B)Wahyu Hendra PrabowoNo ratings yet
- FCPS Part 1 syllabus and book recommendationsDocument2 pagesFCPS Part 1 syllabus and book recommendationsAamir HamaadNo ratings yet
- Goldfinger Human Anatomy For Artists PDFDocument4 pagesGoldfinger Human Anatomy For Artists PDFSamuelNo ratings yet
- Experiment - 1 Rheo Leaf CLS-XDocument2 pagesExperiment - 1 Rheo Leaf CLS-Xsatyajit ojhaNo ratings yet
- NCERT Exemplar For Class 9 Science Chapter 5Document27 pagesNCERT Exemplar For Class 9 Science Chapter 5Vidhan PanwarNo ratings yet
- SPCM Scoring Form Analysis ToolDocument6 pagesSPCM Scoring Form Analysis ToolLaura Vanessa Caballero ArizaNo ratings yet
- DigestionDocument6 pagesDigestionswordmy2523No ratings yet
- Cleavage: Types and PatternsDocument15 pagesCleavage: Types and PatternstrivikramraoNo ratings yet
- Laennecs Cirrhosis PathophysiologyDocument2 pagesLaennecs Cirrhosis PathophysiologyTrixie Al Marie100% (3)
- Poultry Necropsy Manual PDFDocument14 pagesPoultry Necropsy Manual PDFفراس فارس صفاقسNo ratings yet
- Microana Prelim ReviewerDocument17 pagesMicroana Prelim ReviewerjerlerositaNo ratings yet
- Nasopalatine Duct CystDocument4 pagesNasopalatine Duct CystVikneswaran Vîçký100% (1)
- Bone and muscle structure, function, and developmentDocument6 pagesBone and muscle structure, function, and developmentLuna ScorpiusNo ratings yet
- Cerebellum PDFDocument31 pagesCerebellum PDFJonathan PaghubasanNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (14)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisFrom EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisRating: 5 out of 5 stars5/5 (8)