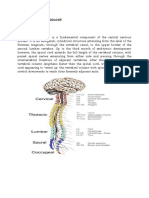
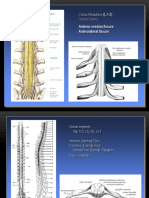
You might also like
- Thoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeFrom EverandThoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeNo ratings yet
- Microsurgical Anatomy of Posterior Fossa VeinsDocument43 pagesMicrosurgical Anatomy of Posterior Fossa VeinsCan EkerNo ratings yet
- Neuroanatomy 11: Brain Ventricles: The Anterior HornDocument3 pagesNeuroanatomy 11: Brain Ventricles: The Anterior HornstuffNo ratings yet
- Central Nervous System: DR Muhammad Ressam NazirDocument28 pagesCentral Nervous System: DR Muhammad Ressam NazirRessam NazirNo ratings yet
- Neuroanatomy 1Document65 pagesNeuroanatomy 1maNo ratings yet
- Ant 325materialDocument13 pagesAnt 325materialDestiny BlissNo ratings yet
- 2-Meninges and SinusesDocument62 pages2-Meninges and SinusesMajd HallakNo ratings yet
- Anatomy of the Ear and its Three RegionsDocument14 pagesAnatomy of the Ear and its Three RegionsBogdan PraščevićNo ratings yet
- Introduction to the Cranial Cavity and its StructuresDocument35 pagesIntroduction to the Cranial Cavity and its StructuresexoNo ratings yet
- Iv. The Spinal Cord: Cordotomy Lumbar PunctureDocument0 pagesIv. The Spinal Cord: Cordotomy Lumbar PunctureAnonymous t5TDwdNo ratings yet
- Tentorium IncisuraDocument24 pagesTentorium IncisuraIndra Hadiandite100% (1)
- The Ventricular SystemDocument6 pagesThe Ventricular SystemM Arsalan TariqNo ratings yet
- Alberta Stroke Program Early CT Score - 1Document4 pagesAlberta Stroke Program Early CT Score - 1Ardina Nur PramudhitaNo ratings yet
- Head and Neck Anatomy GuideDocument16 pagesHead and Neck Anatomy GuideArijeet77No ratings yet
- Pharynx Anatomy - Overview, Gross Anatomy, Microscopic AnatomyDocument9 pagesPharynx Anatomy - Overview, Gross Anatomy, Microscopic AnatomyCarmen-BadeaNo ratings yet
- 3 The ScalpDocument7 pages3 The ScalpAaaNo ratings yet
- 06 Lec 11 12 Anatomy First YearDocument13 pages06 Lec 11 12 Anatomy First YearS:M:ENo ratings yet
- Brainstem and Nuclei Condensed Grayscale SlidesDocument17 pagesBrainstem and Nuclei Condensed Grayscale SlidesMobarobberNo ratings yet
- Spinal CordDocument9 pagesSpinal Cordpromisearinzechi4No ratings yet
- Growth and Development of Cranial Base: SeminarDocument34 pagesGrowth and Development of Cranial Base: SeminarMNSNo ratings yet
- Lateral Ventricles: The Ventricular System of The Human BrainDocument4 pagesLateral Ventricles: The Ventricular System of The Human BrainMarNo ratings yet
- Guide to the Human Brain's Cerebrum and DiencephalonDocument11 pagesGuide to the Human Brain's Cerebrum and DiencephalonDerick JuanNo ratings yet
- Neuroanatomy, Cerebellum: Statpearls (Internet) - Treasure Island (FL) : Statpearls Publishing 2023 JanDocument5 pagesNeuroanatomy, Cerebellum: Statpearls (Internet) - Treasure Island (FL) : Statpearls Publishing 2023 JanANISA RIFKA RIDHONo ratings yet
- Brain and Spinal Cord Anatomy and PhysiologyDocument4 pagesBrain and Spinal Cord Anatomy and PhysiologyynecesityNo ratings yet
- Neuroanatomy, Cerebellar Dysfunction: Gross AnatomyDocument5 pagesNeuroanatomy, Cerebellar Dysfunction: Gross AnatomyANISA RIFKA RIDHONo ratings yet
- Anatomy Physiology of The EarDocument8 pagesAnatomy Physiology of The EargheaastridgayatriNo ratings yet
- ANATOMY AND PHYSIOLOGY OF THE VESTIBULAR ORGAN AND NEURAL PATHWAYSDocument56 pagesANATOMY AND PHYSIOLOGY OF THE VESTIBULAR ORGAN AND NEURAL PATHWAYSVineet Chadha0% (1)
- Ecr2016 C-0757Document28 pagesEcr2016 C-0757mloureiro17No ratings yet
- Head & Neck Blood Vessels Guide - Arteries, Veins & Clinical ImportanceDocument20 pagesHead & Neck Blood Vessels Guide - Arteries, Veins & Clinical ImportancecheckmateNo ratings yet
- Enfermedad Vestibular FelinaDocument8 pagesEnfermedad Vestibular FelinaPamela Susana Rivera GómezNo ratings yet
- Lec 6 Anatomy First StageDocument36 pagesLec 6 Anatomy First Stagespo9equtabaNo ratings yet
- Human Anatomy Assignment 1. DR Onanuga-2021Document7 pagesHuman Anatomy Assignment 1. DR Onanuga-2021AlexanderNo ratings yet
- The Anterior and Middle Cranial BaseDocument30 pagesThe Anterior and Middle Cranial Basejuan valencia sotoNo ratings yet
- General Anatomy Head and Abdomen: Mandibular AngleDocument17 pagesGeneral Anatomy Head and Abdomen: Mandibular AnglesdsddsdsNo ratings yet
- Anatomia y Fisiologia Del DiafragmaDocument11 pagesAnatomia y Fisiologia Del Diafragmaflechudo1235408No ratings yet
- Parvertebral Blokade of TheDocument11 pagesParvertebral Blokade of TheClínica Veterinaria TODOVETNo ratings yet
- Branchial ArchesDocument49 pagesBranchial ArchesSherin ThomasNo ratings yet
- Physio. Psych Structure-Of-The-Nervous-System-Fact-SheetDocument7 pagesPhysio. Psych Structure-Of-The-Nervous-System-Fact-SheetJustynmae JusainNo ratings yet
- SpinalDocument3 pagesSpinalDr.Mahesh kumar GuptaNo ratings yet
- Neuro AnatomyDocument49 pagesNeuro AnatomyShalini SinghNo ratings yet
- Anatomy and Physiology The Spinal CordDocument11 pagesAnatomy and Physiology The Spinal CordKatrina CaveNo ratings yet
- Anatomy and Physiology of The Ear PDFDocument12 pagesAnatomy and Physiology of The Ear PDFDeshini Balasubramaniam83% (6)
- Essay Solution On Space in The SkullDocument21 pagesEssay Solution On Space in The SkullEmmanuel IshiomaNo ratings yet
- Acoustic SensitivityDocument13 pagesAcoustic SensitivityJean Pierre Chastre LuzaNo ratings yet
- Jurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas ADocument17 pagesJurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas AChristopher Muhamad Zildan HNo ratings yet
- CHAPTER 56 The DiaphragmDocument64 pagesCHAPTER 56 The DiaphragmAbeNo ratings yet
- Natomy of Larynx: Moderator: DR - Manas Presenter:Ravindra.DDocument89 pagesNatomy of Larynx: Moderator: DR - Manas Presenter:Ravindra.DVandana RaviNo ratings yet
- Head Lecture NotesDocument49 pagesHead Lecture Notesvipul51190No ratings yet
- Orbital Anatomy: R 2018 Wolters Kluwer Health, Inc. All Rights ReservedDocument19 pagesOrbital Anatomy: R 2018 Wolters Kluwer Health, Inc. All Rights ReservedAna MariaNo ratings yet
- Head &neck 2024Document68 pagesHead &neck 2024danaelmarasy60No ratings yet
- Chronic TympanomastoiditisDocument25 pagesChronic Tympanomastoiditispatriciaberedo100% (1)
- "Root of Neck": Dr. Shoukat Ali MemonDocument31 pages"Root of Neck": Dr. Shoukat Ali MemonRamesh KumarNo ratings yet
- Anatomy of NeckDocument100 pagesAnatomy of NeckdiptidigheNo ratings yet
- Nro NotessafdaadDocument199 pagesNro NotessafdaadMunawwar AwaNo ratings yet
- Attachment 1Document39 pagesAttachment 1wingakira5No ratings yet
- Cavities of The Neuro and ViscerocraniumDocument79 pagesCavities of The Neuro and ViscerocraniumSchönberger AlbertNo ratings yet
- Blood Supply of Spinal Cord... 2022Document31 pagesBlood Supply of Spinal Cord... 2022David Pinto SilvaNo ratings yet
- Aa AnatomyDocument109 pagesAa AnatomyshailendraNo ratings yet
- The Respiratory TractDocument79 pagesThe Respiratory TractbayennNo ratings yet
- Blood Supply of The BrainDocument32 pagesBlood Supply of The BrainShimmering MoonNo ratings yet
- An Update On COVID-19 and Pregnancy: Djamieson@emory - EduDocument10 pagesAn Update On COVID-19 and Pregnancy: Djamieson@emory - EduRaul DoctoNo ratings yet
- 1998 Volvo s70-v70Document52 pages1998 Volvo s70-v70Raul DoctoNo ratings yet
- BMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PDocument13 pagesBMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PRaul DoctoNo ratings yet
- Satoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFDocument44 pagesSatoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFLuisNo ratings yet
- Seminar: Jan Claassen, Soojin ParkDocument17 pagesSeminar: Jan Claassen, Soojin ParkRaul DoctoNo ratings yet
- Central Venous Pressure (CVP)Document3 pagesCentral Venous Pressure (CVP)Raul DoctoNo ratings yet
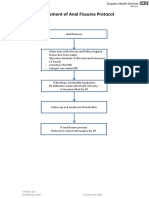
- Anal Fissures PDF 14 Sept-115042Document1 pageAnal Fissures PDF 14 Sept-115042Raul DoctoNo ratings yet
- GINA Coming SoonDocument1 pageGINA Coming SoonRaul DoctoNo ratings yet
- X3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutDocument3 pagesX3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutRaul DoctoNo ratings yet
- 1998 Volvo S70-V70Document52 pages1998 Volvo S70-V70cosminubNo ratings yet
- 2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureDocument167 pages2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureRaul DoctoNo ratings yet
- Ascoping Review of The Pathophysiology of COVID-19Document16 pagesAscoping Review of The Pathophysiology of COVID-19Raul DoctoNo ratings yet
- Rapid Diagnostic Testing For SARS-CoV-2Document9 pagesRapid Diagnostic Testing For SARS-CoV-2Wilfredo NuñezNo ratings yet
- Why Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadDocument15 pagesWhy Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadRaul Docto100% (1)
- Obstetrics & GynecologyDocument10 pagesObstetrics & GynecologyRaul DoctoNo ratings yet
- How To Install Google Play On The Amazon Fire HD 8Document7 pagesHow To Install Google Play On The Amazon Fire HD 8Raul DoctoNo ratings yet
- Robba2021 Article BasicUltrasoundHead-to-toeSkilDocument21 pagesRobba2021 Article BasicUltrasoundHead-to-toeSkilRaul DoctoNo ratings yet
- Complications of HysterectomyDocument17 pagesComplications of HysterectomyRaul DoctoNo ratings yet
- Standardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Document8 pagesStandardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Raul DoctoNo ratings yet
- AJOG Implementation of A Conservative ChecklistDocument9 pagesAJOG Implementation of A Conservative ChecklistRaul DoctoNo ratings yet
- Understanding The Role of Host Response in Influenza PneumonitisDocument3 pagesUnderstanding The Role of Host Response in Influenza PneumonitisRaul DoctoNo ratings yet
- Vlaar2021 Article TransfusionStrategiesInBleedinDocument25 pagesVlaar2021 Article TransfusionStrategiesInBleedinRaul DoctoNo ratings yet
- Case 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaDocument7 pagesCase 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaRaul DoctoNo ratings yet
- Alcohol y Metabolismo HumanoDocument21 pagesAlcohol y Metabolismo HumanoDani BelloNo ratings yet
- Surviving Sepsis Campaign International.21Document81 pagesSurviving Sepsis Campaign International.21jamesteryNo ratings yet
- Tugwell 1985Document13 pagesTugwell 1985Raul DoctoNo ratings yet
- A Framework For Standardized Management of Intrapartum PDFDocument6 pagesA Framework For Standardized Management of Intrapartum PDFRaul A UrbinaNo ratings yet
- 2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisDocument144 pages2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisSara MendesNo ratings yet
- MCU 2021 The Ventilator Book 3rd EditionDocument280 pagesMCU 2021 The Ventilator Book 3rd EditionAlvaro Estupiñan100% (2)
- 2014-08-13 Gynae Positioning Presentation - SlidesDocument14 pages2014-08-13 Gynae Positioning Presentation - SlidesIrina MincuNo ratings yet
- 2011 @radproflib D Karthikeyan Step PDFDocument310 pages2011 @radproflib D Karthikeyan Step PDFToma BichirNo ratings yet
- Atlas of Human Anatomy: Prof. Dr. Med. Petra Köpf-Maier, BerlinDocument14 pagesAtlas of Human Anatomy: Prof. Dr. Med. Petra Köpf-Maier, BerlinEspoire LibertéNo ratings yet
- Pathophysiology of HydrocephalusDocument3 pagesPathophysiology of HydrocephalusJan Rae Barnatia AtienzaNo ratings yet
- Kuliah Medula SpinalisDocument85 pagesKuliah Medula SpinalisMuhamad Thohar ArifinNo ratings yet
- EENT Instillation and IrrigationDocument13 pagesEENT Instillation and Irrigationplebur100% (1)
- Assessment of Tympanic Membrane Perforation in AdultDocument4 pagesAssessment of Tympanic Membrane Perforation in AdultVikrant Vaze100% (1)
- Bickerstaff Brainstem Encephalitis Diagnosis and TreatmentDocument7 pagesBickerstaff Brainstem Encephalitis Diagnosis and TreatmentAubrey Unique EvangelistaNo ratings yet
- Neuroanatomy Brainstem SummaryDocument11 pagesNeuroanatomy Brainstem SummaryStd Dlshsi100% (2)
- Facs Paul Ekman PDFDocument2 pagesFacs Paul Ekman PDFDana0% (2)
- Physiology of Somatosensory CortexDocument48 pagesPhysiology of Somatosensory CortexTeeNo ratings yet
- Facial Feature ExamplesDocument13 pagesFacial Feature ExamplesRick Jackson100% (1)
- ELIGIBLE CANDIDATESDocument962 pagesELIGIBLE CANDIDATESamanvirbNo ratings yet
- Maxillary Sinus Augmentation: Review ArticleDocument6 pagesMaxillary Sinus Augmentation: Review ArticlePriliNo ratings yet
- Deskripsi TemanDocument2 pagesDeskripsi TemanTimoriaty MorindaNo ratings yet
- Kuisioner Gangguan Penghidu - KTI - Iqbal MuhammadDocument3 pagesKuisioner Gangguan Penghidu - KTI - Iqbal MuhammadIqbal Muhammad Yaa-Begitulah100% (1)
- ENT OsceDocument14 pagesENT OsceRj PolvorosaNo ratings yet
- ThyroidectomyDocument2 pagesThyroidectomykzone2290No ratings yet
- Vascular Anatomy and CSF SpacesDocument64 pagesVascular Anatomy and CSF SpacesJha K SujitNo ratings yet
- Blood Supply To The Human Spinal Cord. I. Anatomy and HemodynamicsDocument13 pagesBlood Supply To The Human Spinal Cord. I. Anatomy and HemodynamicsAnusha SebastianNo ratings yet
- Anatomy of the Nose: Bones, Cartilages, Ligaments, Blood SupplyDocument48 pagesAnatomy of the Nose: Bones, Cartilages, Ligaments, Blood SupplyShivaramanRamachandranNo ratings yet
- Postnatal Growth of The Cranial VaultDocument6 pagesPostnatal Growth of The Cranial VaultCornelia GheorghitaNo ratings yet
- Multiple-Choice Questions: Chapter 49 Nervous SystemsDocument10 pagesMultiple-Choice Questions: Chapter 49 Nervous SystemsTrajceNo ratings yet
- Praktikum Endokrin Activity 1 Kelompok 2Document8 pagesPraktikum Endokrin Activity 1 Kelompok 2kuanlins crushNo ratings yet
- Elements of Morphology Human Malformation TerminologyDocument3 pagesElements of Morphology Human Malformation TerminologyWindsurfingFinnNo ratings yet
- Thyroid PowerPoint PresentationDocument31 pagesThyroid PowerPoint PresentationSadashivayya SoppimathNo ratings yet
- Cranial NervesDocument4 pagesCranial NervesClaudio Antonio MutimucuioNo ratings yet
- Fibrous JointDocument5 pagesFibrous JointmamatnamakuNo ratings yet
- 00 H&N NotesDocument16 pages00 H&N NotesHythem HashimNo ratings yet
- Comparison of Graft Uptake and Post Operative Hearing Between Cartilage Rim Augmented Fascia and Temporalis Fascia TympanoplastyDocument7 pagesComparison of Graft Uptake and Post Operative Hearing Between Cartilage Rim Augmented Fascia and Temporalis Fascia TympanoplastyInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Nasopharyngeal Angiofibroma Treatment OptionsDocument51 pagesNasopharyngeal Angiofibroma Treatment OptionsAbhishek ShahNo ratings yet