You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Senate Doc 93-62 Title Acknowledgements IntroDocument23 pagesSenate Doc 93-62 Title Acknowledgements IntroJargons Nether EnNo ratings yet
- Resveratrol and Its Effects On Human Health and LongevityDocument147 pagesResveratrol and Its Effects On Human Health and Longevity9continents100% (1)
- ING - John Eliot - Models of Psychological Space - Psychometric, Developmental, and Experimental Approaches-Springer-Verlag New York (1987) PDFDocument210 pagesING - John Eliot - Models of Psychological Space - Psychometric, Developmental, and Experimental Approaches-Springer-Verlag New York (1987) PDFSofia CardosoNo ratings yet
- DYNSIM Best Practices 1 - CompressorDocument21 pagesDYNSIM Best Practices 1 - CompressorEmre TokgözNo ratings yet
- Bucket Elevator Calculation Rev 1Document8 pagesBucket Elevator Calculation Rev 1Irfan AhmedNo ratings yet
- Lab Report Marcet BoilerDocument8 pagesLab Report Marcet BoilerAnis Thuraiya100% (1)
- 2004 VCF Tables - SGS Implementation (A4)Document1 page2004 VCF Tables - SGS Implementation (A4)Adrian RuizNo ratings yet
- Essays On The Theory of Numbers - Richard DedekindDocument78 pagesEssays On The Theory of Numbers - Richard DedekindJargons Nether EnNo ratings yet
- Graphene Oxide Nanomaterial Exposures On The Marine Bivalve, Crassostrea VirginicaDocument25 pagesGraphene Oxide Nanomaterial Exposures On The Marine Bivalve, Crassostrea VirginicaJargons Nether EnNo ratings yet
- Graphene Oxide Nanosheets For Tumor Cell TherapyDocument23 pagesGraphene Oxide Nanosheets For Tumor Cell TherapyJargons Nether EnNo ratings yet
- 1-Nano-Graphene Oxide As A Nanocarrier For Delivering Mixed Anticancer Drugs To Improve Anticancer ActivityDocument15 pages1-Nano-Graphene Oxide As A Nanocarrier For Delivering Mixed Anticancer Drugs To Improve Anticancer ActivityJargons Nether EnNo ratings yet
- Design and Characterization of Chitosan-Graphene Oxide Nanocomposites For The Delivery of ProanthocyanidinsDocument10 pagesDesign and Characterization of Chitosan-Graphene Oxide Nanocomposites For The Delivery of ProanthocyanidinsJargons Nether EnNo ratings yet
- Essays On The Theory of Numbers - Richard DedekindDocument78 pagesEssays On The Theory of Numbers - Richard DedekindJargons Nether EnNo ratings yet
- The Sars-Cov-2 Delta Variant Is Poised To Acquire Complete Resistance To Wild-Type Spike VaccinesDocument31 pagesThe Sars-Cov-2 Delta Variant Is Poised To Acquire Complete Resistance To Wild-Type Spike VaccinesRainer Yodha ReswaraNo ratings yet
- Medieval Jewish Exegesis of Psalm 2Document22 pagesMedieval Jewish Exegesis of Psalm 2Jargons Nether EnNo ratings yet
- Sub-Acute Toxicity Study of Graphene Oxide in The Sprague-Dawley RatDocument13 pagesSub-Acute Toxicity Study of Graphene Oxide in The Sprague-Dawley RatJargons Nether EnNo ratings yet
- 0-Usability of Graphene Oxide As A Mycotoxin BinderDocument12 pages0-Usability of Graphene Oxide As A Mycotoxin BinderJargons Nether EnNo ratings yet
- Short-Term Inhalation Study of Graphene Oxide NanoplatesDocument28 pagesShort-Term Inhalation Study of Graphene Oxide NanoplatesJargons Nether EnNo ratings yet
- Intestinal Barrier Against Graphene Oxide ToxicityDocument16 pagesIntestinal Barrier Against Graphene Oxide ToxicityJargons Nether EnNo ratings yet
- Graphene-Based Nanomaterials Toxicity in FishDocument37 pagesGraphene-Based Nanomaterials Toxicity in FishJargons Nether EnNo ratings yet
- Toxicity Evaluation of Graphene Oxide in Rat KidneysDocument15 pagesToxicity Evaluation of Graphene Oxide in Rat KidneysJargons Nether EnNo ratings yet
- Toxicity of Graphene Shells, Graphene Oxide, and Graphene Oxide PaperDocument10 pagesToxicity of Graphene Shells, Graphene Oxide, and Graphene Oxide PaperJargons Nether EnNo ratings yet
- Graphene Oxide Nanoparticles Having Long Wavelength Absorbing ChlorinsDocument17 pagesGraphene Oxide Nanoparticles Having Long Wavelength Absorbing ChlorinsJargons Nether EnNo ratings yet
- Toxicity Assessment of Grahphen Oxide and TitaniumDocument7 pagesToxicity Assessment of Grahphen Oxide and TitaniumJargons Nether EnNo ratings yet
- Toxicity Assessment of Graphene Oxide On The Embryonic Development of ZebrafishDocument19 pagesToxicity Assessment of Graphene Oxide On The Embryonic Development of ZebrafishJargons Nether EnNo ratings yet
- Efect of Ingestion of Whey, Casein, or Soy Proteins On Mixed Muscle Protein Synthesis in Resistance ExerciseDocument6 pagesEfect of Ingestion of Whey, Casein, or Soy Proteins On Mixed Muscle Protein Synthesis in Resistance ExerciseJargons Nether EnNo ratings yet
- C Elegans Mir-235 Regulation of Graphene Oxide TixicityDocument14 pagesC Elegans Mir-235 Regulation of Graphene Oxide TixicityJargons Nether EnNo ratings yet
- Graphene Oxide Negatively Regulates Cell Cycle in EmbryoDocument9 pagesGraphene Oxide Negatively Regulates Cell Cycle in EmbryoJargons Nether EnNo ratings yet
- Comparative Toxicity of Graphene Oxide & Zinc OxideDocument14 pagesComparative Toxicity of Graphene Oxide & Zinc OxideJargons Nether EnNo ratings yet
- Toxicity Studies On Graphene-Based NanomaterialsDocument22 pagesToxicity Studies On Graphene-Based NanomaterialsJargons Nether EnNo ratings yet
- The Effect of Fluoride On The Physiology of The Pineal GlandDocument297 pagesThe Effect of Fluoride On The Physiology of The Pineal GlandJargons Nether EnNo ratings yet
- Origins of Boolean Algebra in The Logic of Classes - John Venn and C S Pierce 22-5-2011Document35 pagesOrigins of Boolean Algebra in The Logic of Classes - John Venn and C S Pierce 22-5-2011Jargons Nether EnNo ratings yet
- Graphic Nanomaterial CytotoxicityDocument36 pagesGraphic Nanomaterial CytotoxicityJargons Nether EnNo ratings yet
- Toxicity Assessment of Graphene Oxide On The Embryonic Development of ZebrafishDocument19 pagesToxicity Assessment of Graphene Oxide On The Embryonic Development of ZebrafishJargons Nether EnNo ratings yet
- 1 When You Look Up at The Night SkyDocument2 pages1 When You Look Up at The Night SkyArleen J. LimNo ratings yet
- TDS Byk-1780 enDocument2 pagesTDS Byk-1780 enabhijit.home2022No ratings yet
- Catalogue Havells International CablesDocument76 pagesCatalogue Havells International Cablessiddhant103No ratings yet
- Asme Sec 1 NDTDocument1 pageAsme Sec 1 NDTArul Edwin VijayNo ratings yet
- Power Lines Filters For SPSDocument6 pagesPower Lines Filters For SPSalesysNo ratings yet
- 01 - ED Technical Training Part 1Document76 pages01 - ED Technical Training Part 1Калин АнгеловNo ratings yet
- Yongnuo 560 EX II Flash User ManualDocument1 pageYongnuo 560 EX II Flash User ManualEj CoombsNo ratings yet
- Classical MechanicsDocument4 pagesClassical MechanicsSamantha GreyNo ratings yet
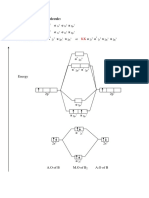
- Molecular Orbital Diagram For B, C, N, O, F and NeDocument12 pagesMolecular Orbital Diagram For B, C, N, O, F and NeGods ArmyNo ratings yet
- Apollon 1Document42 pagesApollon 1Munteanu Maria-MagdalenaNo ratings yet
- Mechanics: Harmonic Oscillation of A String PendulumDocument4 pagesMechanics: Harmonic Oscillation of A String Pendulumfatjonmusli2016100% (1)
- 2 Stress (As of Week 3)Document86 pages2 Stress (As of Week 3)JAMES TRIXIAN PARAJASNo ratings yet
- ST Semi Exam No Answer PDFDocument5 pagesST Semi Exam No Answer PDFBrian UncianoNo ratings yet
- Student Exploration: Doppler Shift: Prior Knowledge Questions (Do These BEFORE Using The GizmoDocument4 pagesStudent Exploration: Doppler Shift: Prior Knowledge Questions (Do These BEFORE Using The GizmoCynta’jah AllisonNo ratings yet
- Astm A320Document7 pagesAstm A320Adán Vera GoNo ratings yet
- Wave OpticsDocument13 pagesWave OpticsBorra ram kumarNo ratings yet
- The Complete Hermes Programme For External Cylindrical GrindingDocument16 pagesThe Complete Hermes Programme For External Cylindrical GrindingFlavioNo ratings yet
- Padeye8t (Without Cheek Plate)Document2 pagesPadeye8t (Without Cheek Plate)Mohd Khir Johari100% (1)
- 09-07-23 JR - Iit Star Co-Sc (Model-A) Jee Adv 2017 (P-II) Wat-11 QP FinalDocument19 pages09-07-23 JR - Iit Star Co-Sc (Model-A) Jee Adv 2017 (P-II) Wat-11 QP FinalRutvika Chowdary NemalapudiNo ratings yet
- CPC 100 User Manual ENUDocument77 pagesCPC 100 User Manual ENUJose Ramirez TorresNo ratings yet
- Lưu ý: - Thí sinh làm bài trên tờ giấy thi, ghi theo đúng thứ tự câu từ 1 đến 100Document6 pagesLưu ý: - Thí sinh làm bài trên tờ giấy thi, ghi theo đúng thứ tự câu từ 1 đến 100Ngọc VươngNo ratings yet
- Introduction To Fluid Mechanics and Properties of FluidsDocument27 pagesIntroduction To Fluid Mechanics and Properties of FluidsAyumi QuiomNo ratings yet
- Mechanical Engineering Hydraulic Mechanics Important MCQ PDFDocument11 pagesMechanical Engineering Hydraulic Mechanics Important MCQ PDFGaneshNo ratings yet
- SVG 203 Intro Remote Sensing-General 2Document117 pagesSVG 203 Intro Remote Sensing-General 2CEO DimejiNo ratings yet
- Extracted Pages From General Chemistry, 6th Ed - BradyDocument9 pagesExtracted Pages From General Chemistry, 6th Ed - BradyAdi NugrahaNo ratings yet