You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- ملزمة دكتور ضياء التدريب الصيفيDocument128 pagesملزمة دكتور ضياء التدريب الصيفيReyam Sadek95100% (2)
- CVB Feed Table 2021 544Document700 pagesCVB Feed Table 2021 544prishix0% (1)
- Perpfn: 1. Composition and Nutrient Determination ValueDocument14 pagesPerpfn: 1. Composition and Nutrient Determination ValueprishixNo ratings yet
- Silsilah Wangsa RajasaDocument1 pageSilsilah Wangsa RajasaprishixNo ratings yet
- Curricullum Vitae Teguh Priatno 7Document5 pagesCurricullum Vitae Teguh Priatno 7prishixNo ratings yet
- COC PremixDocument2 pagesCOC PremixprishixNo ratings yet
- NO Ingredient Per KG Product SatuanDocument2 pagesNO Ingredient Per KG Product SatuanprishixNo ratings yet
- Update Creamino Behn MeyerDocument14 pagesUpdate Creamino Behn Meyerprishix100% (1)
- CTG Analyzer: A Graphical User Interface For CardiotocographyDocument4 pagesCTG Analyzer: A Graphical User Interface For CardiotocographySebastian PerezNo ratings yet
- Womens Self-Management of AsthmaDocument24 pagesWomens Self-Management of AsthmaCenter for Managing Chronic DiseaseNo ratings yet
- Burn GuidelineDocument7 pagesBurn GuidelinegreenNo ratings yet
- Obstructive Sleep Apnea - An Overview - Prof. Siraj WaliDocument95 pagesObstructive Sleep Apnea - An Overview - Prof. Siraj WaliGhaida AlshehriNo ratings yet
- Respiratory System MedicationsDocument2 pagesRespiratory System Medicationsmlbrown8No ratings yet
- LEUKEMIA, MPDs & MDSsDocument11 pagesLEUKEMIA, MPDs & MDSsMary Kaye Yvonne OtillaNo ratings yet
- (English) 5 Daily Habits For Higher Testosterone Levels (DownSub - Com)Document3 pages(English) 5 Daily Habits For Higher Testosterone Levels (DownSub - Com)Anh Văn TrầnNo ratings yet
- Prevalence of Behavioral and Emotional Disorders ADocument7 pagesPrevalence of Behavioral and Emotional Disorders AEricka GutiérrezNo ratings yet
- Vital Signs LectureDocument67 pagesVital Signs LectureJayrelle D. SafranNo ratings yet
- Wellness: Objectives of The Session: HEALTH: WHO, 1947Document21 pagesWellness: Objectives of The Session: HEALTH: WHO, 1947Angelo HuligangaNo ratings yet
- Jurnal 3Document7 pagesJurnal 3Richa FarellaNo ratings yet
- A Toxic Triangle of Destructive Leadership at Bristol Royal InfirmaryDocument19 pagesA Toxic Triangle of Destructive Leadership at Bristol Royal InfirmaryJesicca DeviyantiNo ratings yet
- Content ServerDocument13 pagesContent ServerMairen RamirezNo ratings yet
- Early Orthodontic Treatment of Skeletal Open-Bite MalocclusionDocument7 pagesEarly Orthodontic Treatment of Skeletal Open-Bite MalocclusionMaria SilvaNo ratings yet
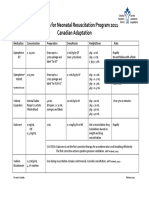
- Medications For Neonatal Resuscitation Program 2011 Canadian AdaptationDocument1 pageMedications For Neonatal Resuscitation Program 2011 Canadian AdaptationrubymayNo ratings yet
- Chemical Burn 3Document4 pagesChemical Burn 3Yeni PuspitasariNo ratings yet
- Self-Resolving Ischemic Third Nerve PalsyDocument10 pagesSelf-Resolving Ischemic Third Nerve PalsyYobbi ArisaputraNo ratings yet
- Class CHAPTER 9 - Circulatory With AnswersDocument59 pagesClass CHAPTER 9 - Circulatory With AnswersSara VinuyaNo ratings yet
- Lithotripsy Salivary Stones OhpDocument5 pagesLithotripsy Salivary Stones Ohpaysha mksNo ratings yet
- Gavin, Nicole - Final Thesis - RedactedDocument245 pagesGavin, Nicole - Final Thesis - RedactedKarlo MedeirosNo ratings yet
- Equipment Devices and Procedures in The Intensive Care Unit PDFDocument16 pagesEquipment Devices and Procedures in The Intensive Care Unit PDFJena Rose ReyesNo ratings yet
- Psychiatric Evaluation of AdultsDocument62 pagesPsychiatric Evaluation of Adultsl1o2stNo ratings yet
- Healthy AgingDocument13 pagesHealthy AgingCarissa Mae Tapec EstradaNo ratings yet
- Outscraper 2024020607591330fd Obstetrician Gynecologist +3Document265 pagesOutscraper 2024020607591330fd Obstetrician Gynecologist +3sanjulata.agriNo ratings yet
- VestibuloplastyDocument57 pagesVestibuloplastyArWiny Wulandari HipiNo ratings yet
- HLTWHS002 PDFDocument4 pagesHLTWHS002 PDFJase Harrison25% (4)
- SchizophreniaDocument32 pagesSchizophreniaHerlina SihalohoNo ratings yet
- GURPS 4e - Injury Template V4Document7 pagesGURPS 4e - Injury Template V4Luca LiperiNo ratings yet
- Morgan Lynn Resume For Weebly OnlyDocument3 pagesMorgan Lynn Resume For Weebly Onlyapi-556602094No ratings yet