You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Presentation by Sana FatimaDocument15 pagesPresentation by Sana FatimaSanaNo ratings yet
- Handling Hazardous Materials at HomeDocument9 pagesHandling Hazardous Materials at HomeKEBAH MORTOLANo ratings yet
- Thesis For MD Community MedicineDocument7 pagesThesis For MD Community Medicinedianaturnerspringfield100% (2)
- The Role of Thiamine Deficiency in Alcoholic Brain DiseaseDocument10 pagesThe Role of Thiamine Deficiency in Alcoholic Brain DiseaseVivekNo ratings yet
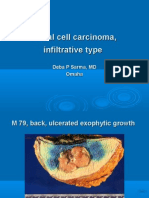
- Basal Cell Carcinoma, Infiltrative TypeDocument8 pagesBasal Cell Carcinoma, Infiltrative TypeDeba P SarmaNo ratings yet
- H Pylori and Peptic UlcersDocument44 pagesH Pylori and Peptic UlcersNdayisaba CorneilleNo ratings yet
- Human Endogenous Retroviruses and AIDS Research: Confusion, Consensus, or Science?Document6 pagesHuman Endogenous Retroviruses and AIDS Research: Confusion, Consensus, or Science?DisicienciaNo ratings yet
- Nursing Practice II - Community Health Nursing and Care of The Mother and Child - RNpediaDocument13 pagesNursing Practice II - Community Health Nursing and Care of The Mother and Child - RNpediaBrianMarBeltranNo ratings yet
- Cause: Lymphatic System Oriental Rat Flea Human Flea Septicemic PlagueDocument3 pagesCause: Lymphatic System Oriental Rat Flea Human Flea Septicemic PlagueMattNo ratings yet
- Screenshot 2021-06-04 at 23.19.29Document72 pagesScreenshot 2021-06-04 at 23.19.29Amina Mohamed AbdikeirNo ratings yet
- We Should Avoid Oily Food . Be Healthy. A. Finally C. Consequently B. in Order To D. So ThatDocument1 pageWe Should Avoid Oily Food . Be Healthy. A. Finally C. Consequently B. in Order To D. So ThathendriNo ratings yet
- Self-Directed Learning (Nur 146 - Clinical Area)Document2 pagesSelf-Directed Learning (Nur 146 - Clinical Area)Dyan Bianca Suaso LastimosaNo ratings yet
- Copd - NCPDocument6 pagesCopd - NCPMonique Sacherow BacherNo ratings yet
- 2022 World Air Quality Report USDocument47 pages2022 World Air Quality Report USJacaranda FM NewsNo ratings yet
- Agent OrangeDocument42 pagesAgent Orangeivandamljanovic007No ratings yet
- 11NCP Ineffective Tissue Perfusion 2018 Revised 3.12.18Document2 pages11NCP Ineffective Tissue Perfusion 2018 Revised 3.12.18Aubrey SungaNo ratings yet
- Kode BPJSDocument40 pagesKode BPJSDwi FortunaNo ratings yet
- Blood GroupsDocument3 pagesBlood GroupsMian Talha KamalNo ratings yet
- Gigantism PresentationDocument24 pagesGigantism PresentationYaseen MohmandNo ratings yet
- PK-225 Tugas 001-I-Pkd Profiling Diri Dinda Olinda DelarosaDocument2 pagesPK-225 Tugas 001-I-Pkd Profiling Diri Dinda Olinda DelarosaIthaPintoNo ratings yet
- English-6-Q2-Module-2-Lesson-2-.version 3 PDFDocument20 pagesEnglish-6-Q2-Module-2-Lesson-2-.version 3 PDFfevilyn limbaga100% (1)
- HARSDocument6 pagesHARSNabila S HusnaNo ratings yet
- Memorial of Team 58Document16 pagesMemorial of Team 58shreyash singhNo ratings yet
- Research Methodology For Business - Healthy LifestyleDocument9 pagesResearch Methodology For Business - Healthy LifestyleRameshwar FundipalleNo ratings yet
- 677 Assignment 1 PDFDocument17 pages677 Assignment 1 PDFinformation pointNo ratings yet
- 1-Embyology Derivatives PhyseoDocument3 pages1-Embyology Derivatives PhyseoGautam ManoharNo ratings yet
- Op-Ed ObesityDocument3 pagesOp-Ed Obesityapi-302568521No ratings yet
- Valencia Colleges (Buk.) Incorporated: NSTP 1: National Service Training Program 1Document6 pagesValencia Colleges (Buk.) Incorporated: NSTP 1: National Service Training Program 1Damon LofrancoNo ratings yet
- Barium EnemaDocument13 pagesBarium EnemaLuisa Manalo100% (1)
- Ebstein Anomaly in The Adult PatientDocument11 pagesEbstein Anomaly in The Adult PatientRJMNo ratings yet