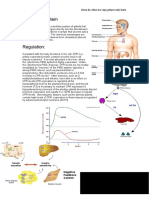
You might also like
- New Insights Into Sympathetic Regulation of Glucose and Fat MetabolismDocument17 pagesNew Insights Into Sympathetic Regulation of Glucose and Fat MetabolismAli HaidarNo ratings yet
- Neuronal Control of Peripheral Insulin Sensitivity and Glucose MetabolismDocument12 pagesNeuronal Control of Peripheral Insulin Sensitivity and Glucose MetabolismValery Alexandra Silva CastellanosNo ratings yet
- Regulation of Hepatic Glucose Metabolism in Health and DiseaseDocument36 pagesRegulation of Hepatic Glucose Metabolism in Health and DiseaseMohammad Hadi SahebiNo ratings yet
- InsulinDocument50 pagesInsulinTandin SonamNo ratings yet
- Endocrine Pancreas FunctionsDocument9 pagesEndocrine Pancreas FunctionsSunardi BahriNo ratings yet
- High-Fat Diet Modulates The Excitability of NeuronDocument19 pagesHigh-Fat Diet Modulates The Excitability of NeuronTuna TunNo ratings yet
- Endocrine SystemDocument2 pagesEndocrine System23aniNo ratings yet
- Introductio 1Document8 pagesIntroductio 1Sikha MohapatraNo ratings yet
- Circadian Rhythms and Diabetes: How Disrupted Clocks Can Lead to DiseaseDocument2 pagesCircadian Rhythms and Diabetes: How Disrupted Clocks Can Lead to DiseaseDwi afrianti Rahma putriNo ratings yet
- Feedback Control MechanismsDocument4 pagesFeedback Control MechanismsJividan SasidranNo ratings yet
- Endo Pancrease ElhDocument27 pagesEndo Pancrease Elhodiodi57No ratings yet
- Psychology of EatingDocument34 pagesPsychology of Eatingtefuture.psychNo ratings yet
- Chapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasDocument17 pagesChapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasGiliana P. García AcevedoNo ratings yet
- Glycolysis - 1 - WUSLDocument52 pagesGlycolysis - 1 - WUSLAruni piyuminiNo ratings yet
- Vasoactive Peptides: ANP, Endothelins, CGRP, and MoreDocument4 pagesVasoactive Peptides: ANP, Endothelins, CGRP, and MoreALNAKINo ratings yet
- Pathogenesis of Delayed Gastric Emptying - UpToDateDocument25 pagesPathogenesis of Delayed Gastric Emptying - UpToDatemayteveronica1000No ratings yet
- Sensors: Glucose Sensing Neurons in The Ventromedial HypothalamusDocument24 pagesSensors: Glucose Sensing Neurons in The Ventromedial HypothalamusNorbert SzilágyiNo ratings yet
- Brain regulation of energy and glucose metabolismDocument12 pagesBrain regulation of energy and glucose metabolismDenise MathreNo ratings yet
- Automatic Nervous SystemDocument68 pagesAutomatic Nervous SystemEurika De GuzmanNo ratings yet
- 08 Autonomic Nervous SystemDocument0 pages08 Autonomic Nervous SystemjuniorebindaNo ratings yet
- Artículo 7. Role of The Gut-Brain Axis in Energy and Glucose MetabolismDocument16 pagesArtículo 7. Role of The Gut-Brain Axis in Energy and Glucose MetabolismJUANITA MESANo ratings yet
- Gastric Emptying DM NEJM 2021Document10 pagesGastric Emptying DM NEJM 2021Luis Alejandro Rosales RenteriaNo ratings yet
- Managing Diabetes and Endocrine DiseasesDocument29 pagesManaging Diabetes and Endocrine DiseasesDesak PratiwiNo ratings yet
- Vegetativen Nerven SistemDocument62 pagesVegetativen Nerven SistemManuela Nikolovska100% (1)
- Insulin and Insulin Resistance in Alzheimer's DiseaseDocument23 pagesInsulin and Insulin Resistance in Alzheimer's DiseaseMarília Cavalcante LemosNo ratings yet
- Pancreatic Regulation of Glucose HomeostasisDocument20 pagesPancreatic Regulation of Glucose Homeostasiseki_herawatiNo ratings yet
- Lecture 14Document25 pagesLecture 14Rezwan Noor NSUNo ratings yet
- FEHM Et Al, 2006Document12 pagesFEHM Et Al, 2006Gilson Mariano NeryNo ratings yet
- PeptidesDocument8 pagesPeptidesSandraosoNo ratings yet
- Principles of Diabetes Mellitus - Ch3FinalDocument16 pagesPrinciples of Diabetes Mellitus - Ch3Finalphoenix sunNo ratings yet
- Abstract (Summary) Full Text: Translatefull Text Turn On Search Term NavigationDocument11 pagesAbstract (Summary) Full Text: Translatefull Text Turn On Search Term NavigationKhairida Hafni LbsNo ratings yet
- Insulinoma in Dogs: A Review: Normal Glucose HomeostasisDocument13 pagesInsulinoma in Dogs: A Review: Normal Glucose HomeostasisBrisbany AlcibarNo ratings yet
- Jais e Bruning (2017) Hypothalamic Inflammation in Obesity and Metabolic DiseaseDocument10 pagesJais e Bruning (2017) Hypothalamic Inflammation in Obesity and Metabolic DiseaseAna Flávia SordiNo ratings yet
- Neuroendocrine Control of Food IntakeDocument11 pagesNeuroendocrine Control of Food IntakeO KiNo ratings yet
- Perspectives in Diabetes: A Critical Role in Metabolic Homeostasis and Disease From C. Elegans To HumansDocument13 pagesPerspectives in Diabetes: A Critical Role in Metabolic Homeostasis and Disease From C. Elegans To Humansetik ainun rohmahNo ratings yet
- Basics in Clinical Nutrition: Carbohydrate Metabolism: Luc TappyDocument4 pagesBasics in Clinical Nutrition: Carbohydrate Metabolism: Luc Tappymarliana100% (1)
- Consequences of Recurrent Hypoglycaemia On Brain Function in DiabetesDocument7 pagesConsequences of Recurrent Hypoglycaemia On Brain Function in DiabetesLuiza KitievaNo ratings yet
- Anatomy, Autonomic Nervous System - StatPearls - NCBI BookshelfDocument10 pagesAnatomy, Autonomic Nervous System - StatPearls - NCBI Bookshelfwood landerNo ratings yet
- Understanding blood glucose regulationDocument3 pagesUnderstanding blood glucose regulationOlive ViaNo ratings yet
- Er 2018-00226Document21 pagesEr 2018-00226Aldo Amed Montaño SalinasNo ratings yet
- Depression, Brain Glucose and Aromatherapy HypothesisDocument10 pagesDepression, Brain Glucose and Aromatherapy HypothesisDani LopezNo ratings yet
- Clinical Manifestations: Psammoma BodiesDocument1 pageClinical Manifestations: Psammoma BodiesshinaNo ratings yet
- Enteric Nervous SystemDocument12 pagesEnteric Nervous SystembaridacheNo ratings yet
- Insulin regulates liver glucose production through both direct and indirect mechanismsDocument9 pagesInsulin regulates liver glucose production through both direct and indirect mechanismsDavid LizardeNo ratings yet
- Type I Diabetes MellitesDocument71 pagesType I Diabetes MellitesMuthu LakshmiNo ratings yet
- Obesity, Metabolism, and Hypertension: M.D. Department BostonDocument9 pagesObesity, Metabolism, and Hypertension: M.D. Department BostonRocio Gutierrez MartinezNo ratings yet
- P'COLOGYDocument18 pagesP'COLOGYHeena BhojwaniNo ratings yet
- tmp24D8 TMPDocument13 pagestmp24D8 TMPFrontiersNo ratings yet
- Insulin Under The Influence of Light: Ursino Gloria, Coppari RobertoDocument11 pagesInsulin Under The Influence of Light: Ursino Gloria, Coppari Robertoro111111No ratings yet
- Regulation of MetabolismDocument42 pagesRegulation of Metabolismامجد حسين جواد كاظمNo ratings yet
- Furness Inervação GIDocument33 pagesFurness Inervação GIAndressa SulamitaNo ratings yet
- Regulation of Visceral Activity4Document54 pagesRegulation of Visceral Activity4Wayne MananaNo ratings yet
- Metabolic Effects of Insulin & Glucagon RegulationDocument33 pagesMetabolic Effects of Insulin & Glucagon RegulationDan Tristam MicabaloNo ratings yet
- Major pathways of carbohydrate metabolismDocument119 pagesMajor pathways of carbohydrate metabolismsimasNo ratings yet
- Role of Insulin in Metabolic Processes (39Document3 pagesRole of Insulin in Metabolic Processes (39Kb AliNo ratings yet
- Phys OkDocument7 pagesPhys OkVivek ChaudharyNo ratings yet
- Autonomics Nervous SystemDocument23 pagesAutonomics Nervous SystemAmarnath SahNo ratings yet
- El EEIDocument9 pagesEl EEIElias Emmanuel JaimeNo ratings yet
- L11 Glucose RegulationDocument21 pagesL11 Glucose RegulationCheng FuNo ratings yet
- Cannabidiol in Anxiety and Sleep - A Large Case SeriesDocument5 pagesCannabidiol in Anxiety and Sleep - A Large Case SeriespantufoNo ratings yet
- Cannabidiol, A Major Non-Psychotropic Cannabis Constituent Enhances Fracture Healing and Stimulates Lysyl..Document10 pagesCannabidiol, A Major Non-Psychotropic Cannabis Constituent Enhances Fracture Healing and Stimulates Lysyl..pantufoNo ratings yet
- Cannabidiol in Anxiety and Sleep - A Large Case SeriesDocument5 pagesCannabidiol in Anxiety and Sleep - A Large Case SeriespantufoNo ratings yet
- Cannabidiol, A Major Non-Psychotropic Cannabis Constituent Enhances Fracture Healing and Stimulates Lysyl..Document10 pagesCannabidiol, A Major Non-Psychotropic Cannabis Constituent Enhances Fracture Healing and Stimulates Lysyl..pantufoNo ratings yet
- Cannabidiol As A Potential Treatment For Anxiety Disorders - Blessing Et Al - 2015Document13 pagesCannabidiol As A Potential Treatment For Anxiety Disorders - Blessing Et Al - 2015Doug OhnemusNo ratings yet
- Cannabidiol and Sports Performance A Narrative RevDocument19 pagesCannabidiol and Sports Performance A Narrative RevpantufoNo ratings yet
- Canabidiol e AtletasDocument8 pagesCanabidiol e AtletaspantufoNo ratings yet
- CBD's effects on inflammation: a reviewDocument10 pagesCBD's effects on inflammation: a reviewpantufoNo ratings yet
- Cannabidiol - From Drug Interaction Potential To Modulation of The GutDocument2 pagesCannabidiol - From Drug Interaction Potential To Modulation of The GutpantufoNo ratings yet
- Cardiac Control: AnthropologyDocument8 pagesCardiac Control: AnthropologypantufoNo ratings yet
- Cannabidiol (CBD) in The Management of Sports-Related TraumaticDocument14 pagesCannabidiol (CBD) in The Management of Sports-Related TraumaticpantufoNo ratings yet
- Cannabidiol and Sports Performance - A Narrative ReviewDocument19 pagesCannabidiol and Sports Performance - A Narrative ReviewpantufoNo ratings yet
- AdjuvantsDocument8 pagesAdjuvantsAnonymous LDivj0jn8qNo ratings yet
- Canabis Intestino Depressao RatosDocument15 pagesCanabis Intestino Depressao RatospantufoNo ratings yet
- CBD's effects on inflammation: a reviewDocument10 pagesCBD's effects on inflammation: a reviewpantufoNo ratings yet
- Clinical Evidence of Magistral Preparations Based On Medicinal CannabisDocument13 pagesClinical Evidence of Magistral Preparations Based On Medicinal CannabispantufoNo ratings yet
- Comparison of Five Oral Cannabidiol Preparations in Adult Humans: Pharmacokinetics, Body Composition, and Heart Rate VariabilityDocument14 pagesComparison of Five Oral Cannabidiol Preparations in Adult Humans: Pharmacokinetics, Body Composition, and Heart Rate VariabilitypantufoNo ratings yet
- Association Between Collagen Production and Mechanical Stretching in Dermal Extracellular MatrixDocument8 pagesAssociation Between Collagen Production and Mechanical Stretching in Dermal Extracellular MatrixpantufoNo ratings yet
- Alzheimer Placebo Controlled StudyDocument13 pagesAlzheimer Placebo Controlled StudypantufoNo ratings yet
- Biofee HRVDocument14 pagesBiofee HRVpantufoNo ratings yet
- The Viability of Microdosing Psychedelics As A Strategy To Enhance Cognition and Well-Being - An Early ReviewDocument11 pagesThe Viability of Microdosing Psychedelics As A Strategy To Enhance Cognition and Well-Being - An Early ReviewpantufoNo ratings yet
- 5157024Document11 pages5157024pantufoNo ratings yet
- Taming of Supernatural Entities and Anim PDFDocument22 pagesTaming of Supernatural Entities and Anim PDFGregorio VelascoNo ratings yet
- Thematic Poster Bioenergetics and Training: (No Relationships Reported)Document2 pagesThematic Poster Bioenergetics and Training: (No Relationships Reported)pantufoNo ratings yet
- Attitude As A Mediator Between Cognitive DissonancDocument10 pagesAttitude As A Mediator Between Cognitive DissonancpantufoNo ratings yet
- Article FluidBalanceInTeamSportAthleteDocument32 pagesArticle FluidBalanceInTeamSportAthletepantufoNo ratings yet
- Photoperiodic Flowering Response of Essential Oil, Grain, and Fiber Hemp (Cannabis Sativa L.) CultivarsDocument22 pagesPhotoperiodic Flowering Response of Essential Oil, Grain, and Fiber Hemp (Cannabis Sativa L.) CultivarspantufoNo ratings yet
- A Warning Against The Negligent Use of CannabidiolDocument5 pagesA Warning Against The Negligent Use of CannabidiolpantufoNo ratings yet
- A Practical Guide To Resonance Frequency Assessment For Heart Rate Variability BiofeedbackDocument14 pagesA Practical Guide To Resonance Frequency Assessment For Heart Rate Variability BiofeedbackpantufoNo ratings yet
- 2020 Article 71597Document9 pages2020 Article 71597pantufoNo ratings yet
- The Role of Medications in Weight LossDocument14 pagesThe Role of Medications in Weight LossWareesa JamshedNo ratings yet
- Obesity TreatmentDocument170 pagesObesity Treatmentojobi joeNo ratings yet
- Focus On SitagliptinDocument90 pagesFocus On SitagliptinBimo Panji KumoroNo ratings yet
- New Practice in Semaglutide On Type-2 Diabetes and Obesity: Clinical Evidence and ExpectationDocument8 pagesNew Practice in Semaglutide On Type-2 Diabetes and Obesity: Clinical Evidence and ExpectationBoris ThielmannNo ratings yet
- Obesity Pathogenesis: Roberto Vettor and Scilla ConciDocument20 pagesObesity Pathogenesis: Roberto Vettor and Scilla ConciErika AvilaNo ratings yet
- Peptide DrugDocument17 pagesPeptide DrugNADIA INDAH FITRIA NINGRUM -100% (1)
- PERKENI (2021) - Konsensus DM Tipe 2Document119 pagesPERKENI (2021) - Konsensus DM Tipe 2Ammalia RachmiNo ratings yet
- GI Long Notes PDFDocument88 pagesGI Long Notes PDFQueenie WongNo ratings yet
- Insulin and IncretinsDocument6 pagesInsulin and IncretinspykkoNo ratings yet
- DrugsDocument98 pagesDrugsReo PratapNo ratings yet
- Farmakologi DMDocument41 pagesFarmakologi DMZainul MuttaqinNo ratings yet
- Antidiabetic Drug PDFDocument91 pagesAntidiabetic Drug PDFAlifia ainushNo ratings yet
- Gut hormones regulating appetiteDocument23 pagesGut hormones regulating appetiteHeba IyadNo ratings yet
- All Types of InsulinDocument18 pagesAll Types of Insulinali mohammedNo ratings yet
- Pedoman DM Tipe 2 2021 - IsbnDocument108 pagesPedoman DM Tipe 2 2021 - IsbnoudyNo ratings yet
- JanubDocument65 pagesJanubRoel PalmairaNo ratings yet
- Report On Januvia DrugDocument10 pagesReport On Januvia DrugyogenaNo ratings yet
- Beta-Cell-Mimetic Designer Cells Provide Closed-Loop Glycemic ControlDocument7 pagesBeta-Cell-Mimetic Designer Cells Provide Closed-Loop Glycemic ControlNando93No ratings yet
- 04-24-2014 Diabetes Drug Classes V0.01asDocument11 pages04-24-2014 Diabetes Drug Classes V0.01asashishhawkingNo ratings yet
- Physician and Patient Perception On Insulin TherapyDocument128 pagesPhysician and Patient Perception On Insulin TherapyEva GabrielNo ratings yet
- Structural Basis For GLP-1 Receptor Activation by LY3502970, An Orally Active Nonpeptide AgonistDocument9 pagesStructural Basis For GLP-1 Receptor Activation by LY3502970, An Orally Active Nonpeptide AgonistClay GranerNo ratings yet
- t2dm Class Comparions ChartDocument1 paget2dm Class Comparions Chartpinkup1199No ratings yet
- Exenatide Response in An Adolescent's Morbid Obesity: Pharmacology & Pharmacy, 2014, 5, 419-424Document6 pagesExenatide Response in An Adolescent's Morbid Obesity: Pharmacology & Pharmacy, 2014, 5, 419-424CamiNo ratings yet
- Case Study 2Document29 pagesCase Study 2drnasim20088171No ratings yet
- Hyperglicemia in Stroke TreatmentDocument13 pagesHyperglicemia in Stroke TreatmentChiquitaPutriVaniaRauNo ratings yet
- Lecture 7 Diabetes Mellitus 2021Document39 pagesLecture 7 Diabetes Mellitus 2021Ahmed ShakerNo ratings yet
- PDF 6Document8 pagesPDF 6Royyan AdiwijayaNo ratings yet
- Tradjenta: Generic Name: (LIN A GLIP Tin) Brand Names: TradjentaDocument5 pagesTradjenta: Generic Name: (LIN A GLIP Tin) Brand Names: TradjentaVidya ShankarNo ratings yet
- APPETITE: Physiological and Neurobiological AspectsDocument8 pagesAPPETITE: Physiological and Neurobiological AspectsTamara Souza RossiNo ratings yet
- Pharmacodynamics of Antidiabetic DrugsDocument25 pagesPharmacodynamics of Antidiabetic DrugsAbdurahman RidhoduaNo ratings yet