You might also like
- The Book of Caterpillars: A Life-Size Guide to Six Hundred Species from Around the WorldFrom EverandThe Book of Caterpillars: A Life-Size Guide to Six Hundred Species from Around the WorldDavid G. JamesRating: 4.5 out of 5 stars4.5/5 (4)
- Out of Africa: Origins of The Taenia Tapeworms in HumansDocument8 pagesOut of Africa: Origins of The Taenia Tapeworms in Humansth3h4ck3rNo ratings yet
- Morphology and Phylogenetic Position of An Unusual Stentor Polymorphus (Ciliophora: Heterotrichea) Without Symbiotic AlgaeDocument8 pagesMorphology and Phylogenetic Position of An Unusual Stentor Polymorphus (Ciliophora: Heterotrichea) Without Symbiotic AlgaeSyifa SariNo ratings yet
- A Taxonomic and Phylogenetic Revision of Penicillium Sec - 2014 - Studies in MycDocument79 pagesA Taxonomic and Phylogenetic Revision of Penicillium Sec - 2014 - Studies in MycUwU SolidustNo ratings yet
- Hernaez Et Al 2010Document8 pagesHernaez Et Al 2010Betel Martinez GuerreroNo ratings yet
- How a Tiny Parasite Controls BlowfliesDocument22 pagesHow a Tiny Parasite Controls BlowfliesjohnNo ratings yet
- Fungal Systematics and Evolution: Symmetrospora (Pucciniomycotina, Cystobasidiomycetes)Document10 pagesFungal Systematics and Evolution: Symmetrospora (Pucciniomycotina, Cystobasidiomycetes)Pham tuan anhNo ratings yet
- TMP BB8 DDocument13 pagesTMP BB8 DFrontiersNo ratings yet
- FasciolaDocument9 pagesFasciolaLakshmiNarayana Reddy BynagariNo ratings yet
- Karyotype and Meiosis Analysis of Four Species of Cameroonian Pyrgomorphidae (Orthoptera)Document9 pagesKaryotype and Meiosis Analysis of Four Species of Cameroonian Pyrgomorphidae (Orthoptera)naufal umerNo ratings yet
- Molecular Investigation and Phylogeny of Anaplasma Spp. in Mediterranean Ruminants Reveal The Presence of Neutrophil-Tropic Strains Closely Related To A. PlatysDocument10 pagesMolecular Investigation and Phylogeny of Anaplasma Spp. in Mediterranean Ruminants Reveal The Presence of Neutrophil-Tropic Strains Closely Related To A. Platysyudhi arjentiniaNo ratings yet
- Hormonios SerpentesDocument18 pagesHormonios SerpentesCamila MoraesNo ratings yet
- JCB 0720Document10 pagesJCB 0720yeetus GGNo ratings yet
- Polystomoides Nelsoni Sp. N. Ventral View of HoloDocument1 pagePolystomoides Nelsoni Sp. N. Ventral View of Holovalerialisseth12No ratings yet
- One Subspecies of The (Gallus Gallus Gallus) : JunglefowlDocument5 pagesOne Subspecies of The (Gallus Gallus Gallus) : JunglefowlCayo GomesNo ratings yet
- Adamowicz Hebert Marinone 2004 Species Diversity DaphniaDocument35 pagesAdamowicz Hebert Marinone 2004 Species Diversity DaphniaDavid CarrozzoNo ratings yet
- Lepto Spiros IsDocument15 pagesLepto Spiros IsShishir AryalNo ratings yet
- Ajol File Journals - 413 - Articles - 61510 - Submission - Proof - 61510 4921 115547 1 10 20101101Document3 pagesAjol File Journals - 413 - Articles - 61510 - Submission - Proof - 61510 4921 115547 1 10 20101101Raul Hernandez SotoNo ratings yet
- Exploring Neotopical Anuran Parasites - A Morphological Life Cycle and Phylogenetic Study of Catadiscus MarinholutziDocument11 pagesExploring Neotopical Anuran Parasites - A Morphological Life Cycle and Phylogenetic Study of Catadiscus MarinholutziMuriloQueirozNo ratings yet
- A Pluralistic Approach in The Study of Pleurotus Species With Emphasis On Compatibility and Physiology of The European MorphotaxaDocument15 pagesA Pluralistic Approach in The Study of Pleurotus Species With Emphasis On Compatibility and Physiology of The European MorphotaxajakoknoxNo ratings yet
- Phyllomedusa 14Document8 pagesPhyllomedusa 14Lendy MilagrosNo ratings yet
- Nematoide Capilaridae Na Língua de Cerdocyon Thous de Vida Livre No BrasilDocument14 pagesNematoide Capilaridae Na Língua de Cerdocyon Thous de Vida Livre No BrasilMaria EduardaNo ratings yet
- Abzhanov Kaufman 2000 EvolutionDocument14 pagesAbzhanov Kaufman 2000 EvolutionDavid CarrozzoNo ratings yet
- International Journal of Acarology: Gaeolaelaps Scarites Sp. Nov., A New Laelapid MiteDocument7 pagesInternational Journal of Acarology: Gaeolaelaps Scarites Sp. Nov., A New Laelapid MiteRani Rahmawati RNo ratings yet
- Harmful Mesostigmatic Mites Ectoparasitic to Honey Bees: Ν.G. Emmanouel, C D - Pelekassis And L.A. SantasDocument7 pagesHarmful Mesostigmatic Mites Ectoparasitic to Honey Bees: Ν.G. Emmanouel, C D - Pelekassis And L.A. SantasΝικόλαος ΚυριαζήςNo ratings yet
- Research Note / Nota Científica: Haematopinus Tuberculatus Infestation and Distribution inDocument6 pagesResearch Note / Nota Científica: Haematopinus Tuberculatus Infestation and Distribution inNanang AdraverycildNo ratings yet
- Studies On Biology and Reproduction of Butterflies (Family: Papilionidae) in Nilgiris Hills, Southern Western Ghats, IndiaDocument11 pagesStudies On Biology and Reproduction of Butterflies (Family: Papilionidae) in Nilgiris Hills, Southern Western Ghats, IndiaEman SamirNo ratings yet
- Pentastomids infect 59% of Australian tropical snakesDocument12 pagesPentastomids infect 59% of Australian tropical snakesEstefania MoralesNo ratings yet
- MATA 2004 ADocument17 pagesMATA 2004 Ajose luisNo ratings yet
- Giardia and GiardiasisDocument16 pagesGiardia and Giardiasisayu05No ratings yet
- Additional Novel Cryptosporidium Genotypes in Orname - 2012 - Veterinary ParasitDocument5 pagesAdditional Novel Cryptosporidium Genotypes in Orname - 2012 - Veterinary Parasitfredys seguraNo ratings yet
- Marine Ecology - 2011 - Cecere - Vegetative Reproduction by Multicellular Propagules in Rhodophyta An OverviewDocument19 pagesMarine Ecology - 2011 - Cecere - Vegetative Reproduction by Multicellular Propagules in Rhodophyta An OverviewKathya Gómez AmigoNo ratings yet
- 782-Texto Del Artículo-2810-1-10-20200830Document16 pages782-Texto Del Artículo-2810-1-10-20200830The BatmanNo ratings yet
- Mite parasitism affects body condition and social structure of colonial batsDocument8 pagesMite parasitism affects body condition and social structure of colonial batsfaradisa35No ratings yet
- Schmidt2004 1Document13 pagesSchmidt2004 1bumble blick crumplestickNo ratings yet
- Weinbeer - Kalko. 2004. Morphological Characterictics Predict Alternate Foraging Strategy and Microhabitat Selection in The Orange-Bellied BatDocument8 pagesWeinbeer - Kalko. 2004. Morphological Characterictics Predict Alternate Foraging Strategy and Microhabitat Selection in The Orange-Bellied BatJ E Fernando CernaNo ratings yet
- Additional Information On Reproductive Behavior of The Red-Breasted Toucan, (Aves: Piciformes: Ramphastidae)Document8 pagesAdditional Information On Reproductive Behavior of The Red-Breasted Toucan, (Aves: Piciformes: Ramphastidae)Miguel ParejoNo ratings yet
- Sehgal 2005Document12 pagesSehgal 2005Gustavo AndresNo ratings yet
- Jurnal Internasional t3b 2Document9 pagesJurnal Internasional t3b 2nanda anisaNo ratings yet
- The Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiteDocument11 pagesThe Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiterachiiidaNo ratings yet
- Ijp: Parasites and Wildlife: SciencedirectDocument6 pagesIjp: Parasites and Wildlife: SciencedirectFitri AnggraeniNo ratings yet
- Wasko 2010Document12 pagesWasko 2010NacorNo ratings yet
- Marshall and Berbee 2010 MBEDocument13 pagesMarshall and Berbee 2010 MBEWyth MarshallNo ratings yet
- MCP 049Document11 pagesMCP 049rachiiidaNo ratings yet
- Guia Morfologia Heces BrasilDocument24 pagesGuia Morfologia Heces BrasilTom BombadilNo ratings yet
- Clave Especies Aspergillus - Samson Et Al 2010Document6 pagesClave Especies Aspergillus - Samson Et Al 2010Liz TrujilloNo ratings yet
- Trematodes (Flukes)Document6 pagesTrematodes (Flukes)COLLEEN GULLENNo ratings yet
- Genetic Determination of The Queen Caste in An Ant Hybrid ZoneDocument4 pagesGenetic Determination of The Queen Caste in An Ant Hybrid ZoneTeresaPeraltaCalvilloNo ratings yet
- Ectoparasites of Bats from Gomantong Forest Reserve in SabahDocument13 pagesEctoparasites of Bats from Gomantong Forest Reserve in SabahEhsan HadiNo ratings yet
- Explosive Breeding Reduces Egg and Tadpole Cannibalism in The Wood FrogDocument9 pagesExplosive Breeding Reduces Egg and Tadpole Cannibalism in The Wood FrogCarolina Gomez SernaNo ratings yet
- 10.1055@s 2006 924177Document8 pages10.1055@s 2006 924177sayyed bassir ajellehNo ratings yet
- EPN Species Found for First Time in JordanDocument3 pagesEPN Species Found for First Time in JordanfadwaeNo ratings yet
- 10 1016@j Actatropica 2006 05 004 PDFDocument3 pages10 1016@j Actatropica 2006 05 004 PDFdianNo ratings yet
- Dun Well 2010Document48 pagesDun Well 2010lacisagNo ratings yet
- Rangel Santos 2009 JParasitol 95 5 1233-1236Document4 pagesRangel Santos 2009 JParasitol 95 5 1233-1236totilas2112No ratings yet
- Wallach Epictia PaperDocument160 pagesWallach Epictia PaperherpmxNo ratings yet
- Phylogenetic Position of Symbiodinium DiDocument9 pagesPhylogenetic Position of Symbiodinium DiThu Anh NguyenNo ratings yet
- Paper 4Document12 pagesPaper 4FarzaneKzrnNo ratings yet
- s10267 007 0364 6Document8 pagess10267 007 0364 6indra maulanaNo ratings yet
- The Harvest of Antaresia Maculosa (Pythonidae) From West Papua, IndonesiaDocument13 pagesThe Harvest of Antaresia Maculosa (Pythonidae) From West Papua, IndonesiaOman SetiyantoNo ratings yet
- Reptiles NYSDEC Environmental Education: For Students in Grades 1 Through 4Document4 pagesReptiles NYSDEC Environmental Education: For Students in Grades 1 Through 4Oman SetiyantoNo ratings yet
- HAYATI Journal of Biosciences: Ida Ayu Ari Janiawati, Mirza Dikari Kusrini, Ani MardiastutiDocument7 pagesHAYATI Journal of Biosciences: Ida Ayu Ari Janiawati, Mirza Dikari Kusrini, Ani MardiastutiOman SetiyantoNo ratings yet
- Pathology of Circulatory DisturbancesDocument72 pagesPathology of Circulatory DisturbancesOman SetiyantoNo ratings yet
- Kuliah Patologi Gangguan Metabolisme 2017Document89 pagesKuliah Patologi Gangguan Metabolisme 2017Oman SetiyantoNo ratings yet
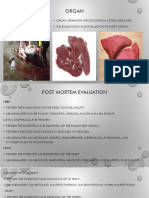
- Organ: - Organ Separation Processe From Cattle Carcasses - The Examination and Evaluation of Every OrganDocument3 pagesOrgan: - Organ Separation Processe From Cattle Carcasses - The Examination and Evaluation of Every OrganOman SetiyantoNo ratings yet
- Vetsminar #6 - Dr. Simon - Heat InjuryDocument34 pagesVetsminar #6 - Dr. Simon - Heat InjuryOman SetiyantoNo ratings yet
- Cellular Injury Necrosis ApoptosisDocument43 pagesCellular Injury Necrosis ApoptosisOman SetiyantoNo ratings yet
- MakalahDocument102 pagesMakalahOman SetiyantoNo ratings yet
- USDA Preharvest Activity SummaryDocument18 pagesUSDA Preharvest Activity SummaryOman SetiyantoNo ratings yet
- Pengaruh Pemberian Chitosan Terhadap Kadar Glukosa Darah Dan Histologi Pankreas Tikus SpragueDocument12 pagesPengaruh Pemberian Chitosan Terhadap Kadar Glukosa Darah Dan Histologi Pankreas Tikus SpragueBilal SabikhismaNo ratings yet
- 2011 MarDocument152 pages2011 MarOman SetiyantoNo ratings yet
- Ipi 31761Document11 pagesIpi 31761Oman SetiyantoNo ratings yet
- Prof. DR - Herry Agoes Hermadi Faculty of Veterinary Medicine Airlangga University SurabayaDocument55 pagesProf. DR - Herry Agoes Hermadi Faculty of Veterinary Medicine Airlangga University SurabayaOman SetiyantoNo ratings yet
- 5 Minute Veterinary Consult Canine and FelineDocument1,016 pages5 Minute Veterinary Consult Canine and FelineMallory Bernstein100% (16)
- Ciri Ciri Fisik Telur Tetas Itik MandalungDocument5 pagesCiri Ciri Fisik Telur Tetas Itik MandalungOman SetiyantoNo ratings yet
- HeterakisDocument6 pagesHeterakisOman SetiyantoNo ratings yet
- 2011 MarDocument152 pages2011 MarOman SetiyantoNo ratings yet
- A Study On The Diversity and Abundance of Spiders in Arakulam VillageDocument5 pagesA Study On The Diversity and Abundance of Spiders in Arakulam Villagefebi chackoNo ratings yet
- AS Biology Unit 2 Topic 4 Plant Structure Biodiversity ConservationDocument4 pagesAS Biology Unit 2 Topic 4 Plant Structure Biodiversity ConservationMilka RahmanNo ratings yet
- Alice in The Land of PlantsDocument384 pagesAlice in The Land of PlantsWellington FavaNo ratings yet
- Guidelines For Biodiversity ResearchDocument6 pagesGuidelines For Biodiversity ResearchAliyyah AndersonNo ratings yet
- Communities WorksheetDocument2 pagesCommunities WorksheetJamie MakrisNo ratings yet
- Maintaining Biodiversity in North-east IndiaDocument10 pagesMaintaining Biodiversity in North-east Indiadr ishani chakrabarttyNo ratings yet
- Systematic BotanyDocument358 pagesSystematic Botanyrandel100% (3)
- Taxonomy and SystematicsDocument22 pagesTaxonomy and SystematicsHyun JiNo ratings yet
- Chapman 2009 NumbersofLivingSpecies Oz World DEHDocument85 pagesChapman 2009 NumbersofLivingSpecies Oz World DEHvero adaroNo ratings yet
- SCIENCE8 Species DiversityDocument8 pagesSCIENCE8 Species DiversityJene kristel ManabatNo ratings yet
- Fish TaxonomyDocument28 pagesFish TaxonomyAntonette OngNo ratings yet
- Dwnload Full Essentials of Physical Anthropology 3rd Edition Clark Test Bank PDFDocument10 pagesDwnload Full Essentials of Physical Anthropology 3rd Edition Clark Test Bank PDFmollybrevet.9xj4b0100% (10)
- Identification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingDocument12 pagesIdentification of Sex-Specific Molecular Markers Using Restriction Site-Associated DNA SequencingFernando Castro EchavezNo ratings yet
- Chap 22Document42 pagesChap 22Parham ElahiNo ratings yet
- Bacillaria Book 2005 PDFDocument135 pagesBacillaria Book 2005 PDFAngie PradaNo ratings yet
- Biodiversity Within A CommunityDocument9 pagesBiodiversity Within A CommunityLibibe IsmaNo ratings yet
- Spiritual Race Aspect ThinkersDocument8 pagesSpiritual Race Aspect Thinkers'рђави ЧаваоNo ratings yet
- The Domestic Dog Its Evolution Behavior and Interactions With People 2ndDocument62 pagesThe Domestic Dog Its Evolution Behavior and Interactions With People 2ndharold.giddings892100% (37)
- Designer Babies Lesson PlanDocument8 pagesDesigner Babies Lesson Planjfeur001100% (1)
- Tolypothrix DistortaDocument13 pagesTolypothrix DistortaIrinaSalariNo ratings yet
- ProposalDocument21 pagesProposalKyle PeñarroyoNo ratings yet
- Hooper Et Al 2005Document33 pagesHooper Et Al 2005Jhonatan Gutierrez100% (1)
- Pseudomonas KoreensisDocument7 pagesPseudomonas KoreensisjmoteizaNo ratings yet
- Scheme of Work Science Stage 7.v1Document44 pagesScheme of Work Science Stage 7.v1Swati Rathod71% (7)
- I. Objectives: The Process of EvolutionDocument3 pagesI. Objectives: The Process of EvolutionEvangelene Esquillo SanaNo ratings yet
- ap-biology-2020-practice-exam-and-notes-3-mcq-answersDocument32 pagesap-biology-2020-practice-exam-and-notes-3-mcq-answersEric ChiangNo ratings yet
- Point of View: Syst. Biol. 69 (6) :1231-1253, 2020Document23 pagesPoint of View: Syst. Biol. 69 (6) :1231-1253, 2020arisNo ratings yet
- 15 Beaks of FinchesDocument8 pages15 Beaks of FinchesKevin LiNo ratings yet
- Comparing DNA Sequences To Understand Evolutionary Relationships With BlastDocument3 pagesComparing DNA Sequences To Understand Evolutionary Relationships With Blastflyawayxx13No ratings yet
- Xi Neet BiologyDocument23 pagesXi Neet BiologyAnonymous eWMnRr70q94% (16)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet