You might also like
- BOSH For SO1 Student Manual SampleDocument65 pagesBOSH For SO1 Student Manual SampleFroilan olazo100% (1)
- Resusciation Equipments in IcuDocument29 pagesResusciation Equipments in Icuvinoli100% (1)
- Medical Microbiology Examination (140 Questions) : First Aid For The Basic Sciences General PrinciplesDocument58 pagesMedical Microbiology Examination (140 Questions) : First Aid For The Basic Sciences General Principlesyashwanth88100% (1)
- Procedure Checklist For Suctioning NCM 112 SL Done Not DoneDocument4 pagesProcedure Checklist For Suctioning NCM 112 SL Done Not DoneIrish Eunice Felix100% (1)
- NP1 - ToprankDocument16 pagesNP1 - ToprankAllaiza Cristille100% (1)
- Ectopic Pregnancy Nursing Care PlansDocument27 pagesEctopic Pregnancy Nursing Care Plansviper7967880% (20)
- DNA Markers for PVY ResistanceDocument10 pagesDNA Markers for PVY ResistanceAnonymous C6laqkPAAKNo ratings yet
- 03 MalenicaDocument9 pages03 MalenicaFilip MartinovićNo ratings yet
- Gobatto 2014Document10 pagesGobatto 2014Francisca Beltrán GuzmánNo ratings yet
- Occurrence and Detection of Sweet Potato Virus DiseaseDocument4 pagesOccurrence and Detection of Sweet Potato Virus DiseaseShailendra RajanNo ratings yet
- Ijab 2015Document4 pagesIjab 2015saadiaNo ratings yet
- s00705 019 04381 ZDocument6 pagess00705 019 04381 ZLaura Paola LópezNo ratings yet
- Donovan2020 Article FirstReportOfCitrusExocortisViDocument3 pagesDonovan2020 Article FirstReportOfCitrusExocortisViimenNo ratings yet
- EJAR - Volume 101 - Issue 3 - Pages 739-750Document12 pagesEJAR - Volume 101 - Issue 3 - Pages 739-750nailulNo ratings yet
- Transmission of MWMV by Aphids 2016Document6 pagesTransmission of MWMV by Aphids 2016Inma Espinar VergelNo ratings yet
- Tmp289a TMPDocument4 pagesTmp289a TMPFrontiersNo ratings yet
- PD-90-1293 Cafrune Et Al 2006Document4 pagesPD-90-1293 Cafrune Et Al 2006Eva CafruneNo ratings yet
- Rice Tungro DiseaseDocument6 pagesRice Tungro DiseaseilmanhasifiNo ratings yet
- PD 90 1004Document8 pagesPD 90 1004bayamcrispyyNo ratings yet
- Amplifi Cation of Emerging Viruses in A Bat ColonyDocument8 pagesAmplifi Cation of Emerging Viruses in A Bat Colonypopayonutz22No ratings yet
- Krzysztof Treder, Włodzimierz Przewodowski, Agnieszka BarnykDocument10 pagesKrzysztof Treder, Włodzimierz Przewodowski, Agnieszka BarnykKrzysztof TrederNo ratings yet
- Genetic Mapping of RYMV Resistance and Relationship to Rice MorphologyDocument14 pagesGenetic Mapping of RYMV Resistance and Relationship to Rice MorphologyCleaver BrightNo ratings yet
- Overview of The PVY Management in France by The Seed Potato Sector (PowerPoint)Document25 pagesOverview of The PVY Management in France by The Seed Potato Sector (PowerPoint)Jorge Luis Alonso G.No ratings yet
- Viruses 11 00045Document25 pagesViruses 11 00045Syarifah TasharaNo ratings yet
- Molecular Detection of Nepoviruses in Pakistani Grapevine Germplasm Using Real Time QRT-PCR and Conventional RT-PCRDocument8 pagesMolecular Detection of Nepoviruses in Pakistani Grapevine Germplasm Using Real Time QRT-PCR and Conventional RT-PCRAisha YounasNo ratings yet
- 1 s2.0 S0168170222003343 MainDocument8 pages1 s2.0 S0168170222003343 MainLaura Paola LópezNo ratings yet
- Box PCRDocument10 pagesBox PCRbabahali7866No ratings yet
- Molecular and Biological Properties of A Putative Partitivirus From Jalapeño Pepper (Capsicum Annuum L.)Document6 pagesMolecular and Biological Properties of A Putative Partitivirus From Jalapeño Pepper (Capsicum Annuum L.)Pamela AboytesNo ratings yet
- Sorho - 2014 - Identification of Viral Disease of Solanaceus in CIDocument8 pagesSorho - 2014 - Identification of Viral Disease of Solanaceus in CIDio DembeleNo ratings yet
- New Mealybug Species Vectoring Grapevine Leafroll-AssociatedDocument7 pagesNew Mealybug Species Vectoring Grapevine Leafroll-Associatedbrkica2011No ratings yet
- Incidence of Viruses in Highbush Blueberry in SerbiaDocument6 pagesIncidence of Viruses in Highbush Blueberry in Serbiakontiki500No ratings yet
- Glomus Bradyrhizobium JaponicumDocument5 pagesGlomus Bradyrhizobium JaponicumFábio Luís MostassoNo ratings yet
- Emma - Pdis 06 22 1350 SCDocument3 pagesEmma - Pdis 06 22 1350 SCOlajide Emmanuel OlorunfemiNo ratings yet
- Grieco 2002Document4 pagesGrieco 2002Camila GamboaNo ratings yet
- Journal Pone 0255029Document12 pagesJournal Pone 0255029Ajzm CompanyNo ratings yet
- EuropaDocument8 pagesEuropaViodelis CrespoNo ratings yet
- Screening of F1 Hybrids and Their Parents For Resistance To Multiple Viruses in Chilli (Capsicum Annuum L.)Document8 pagesScreening of F1 Hybrids and Their Parents For Resistance To Multiple Viruses in Chilli (Capsicum Annuum L.)TJPRC PublicationsNo ratings yet
- Barbier Et Al 2003 SalsomaggioreDocument4 pagesBarbier Et Al 2003 SalsomaggioreennescribeNo ratings yet
- 4 04SNSVActaHortFRESADocument5 pages4 04SNSVActaHortFRESAMartin MorenoNo ratings yet
- in Vitro Antiherpetic Activity of An Aqueous Extract From The Plant Phyllanthus OrbicularisDocument7 pagesin Vitro Antiherpetic Activity of An Aqueous Extract From The Plant Phyllanthus OrbicularisLednys Palomino ZambranoNo ratings yet
- Sub Angui en Pasto GolfDocument6 pagesSub Angui en Pasto GolfEdgar Medina GomezNo ratings yet
- Article ScreeningpepinoDocument14 pagesArticle ScreeningpepinoJuan PachecoNo ratings yet
- Grupo 6Document17 pagesGrupo 6yasminNo ratings yet
- Chapitre SVCDocument18 pagesChapitre SVCAmira BaihaniNo ratings yet
- SMYEPV ProtocolDocument1 pageSMYEPV ProtocolArmin VukojevicNo ratings yet
- 378 FullDocument10 pages378 FullHedi SusantoNo ratings yet
- Meena 2019Document7 pagesMeena 2019PANKAJ YADAVNo ratings yet
- Biological Management of Groundnut Stemrot Pathogen: Sclerotium Rolfsii (Sacc.) by TrichodermavirideDocument8 pagesBiological Management of Groundnut Stemrot Pathogen: Sclerotium Rolfsii (Sacc.) by TrichodermavirideTJPRC PublicationsNo ratings yet
- Early Embryo Invasion As A Determinant in Pea of The Seed Transmission of Pea Seed-Borne Mosaic VirusDocument6 pagesEarly Embryo Invasion As A Determinant in Pea of The Seed Transmission of Pea Seed-Borne Mosaic VirusDenih WahyudinNo ratings yet
- Dialnet ReviewEliminationOfVirusesInPlants 4183676 PDFDocument16 pagesDialnet ReviewEliminationOfVirusesInPlants 4183676 PDFAngelCórdovaNo ratings yet
- DR Dalia LotfyDocument4 pagesDR Dalia LotfyLawangin Khan yousafxaiNo ratings yet
- Lvarez Et Al-2018-Indian Journal of MicrobiologyDocument5 pagesLvarez Et Al-2018-Indian Journal of MicrobiologyUrias GCNo ratings yet
- SPREAD OF TOMATO VIRUSES TO NEW AREA IN PORTUGALDocument2 pagesSPREAD OF TOMATO VIRUSES TO NEW AREA IN PORTUGALjoeNo ratings yet
- Ann Rev of Virology IPNVDocument30 pagesAnn Rev of Virology IPNVRodolfo VelazcoNo ratings yet
- Olesen - 2011 - Multispectral Imaging As A Potential Tool For Seed Health TestingDocument11 pagesOlesen - 2011 - Multispectral Imaging As A Potential Tool For Seed Health TestingtinNo ratings yet
- Prevalence and Genetic Diversity of Human Astroviruses in MexicanDocument7 pagesPrevalence and Genetic Diversity of Human Astroviruses in MexicanGuadalupE Ortiz LópezNo ratings yet
- A Geminivirus-Related DNA Mycovirus That Confers Hypovirulence To A Plant Pathogenic FungusDocument6 pagesA Geminivirus-Related DNA Mycovirus That Confers Hypovirulence To A Plant Pathogenic FungusBeatriz BastidasNo ratings yet
- 2022, Plant, HyperVirulence, PsV5, Puccinia, Agrobacterium-Mediated Transformation AND Plasmid Transformation by PEG - FusariumDocument14 pages2022, Plant, HyperVirulence, PsV5, Puccinia, Agrobacterium-Mediated Transformation AND Plasmid Transformation by PEG - FusariumGorae DollNo ratings yet
- pm7 024 5 enDocument72 pagespm7 024 5 enr8hx4ct6ndNo ratings yet
- Deteksi Dini RTBV Pada Beberapa Generasi F2 Padi Melalui Uji PCRDocument9 pagesDeteksi Dini RTBV Pada Beberapa Generasi F2 Padi Melalui Uji PCRmememeghaNo ratings yet
- Legume-Nodulating Bacteria from Tunisian PasturesDocument8 pagesLegume-Nodulating Bacteria from Tunisian PasturesSAMI DHAOUINo ratings yet
- Synopsis For BSC 2nd DraftDocument11 pagesSynopsis For BSC 2nd DraftArbab KhanNo ratings yet
- Modification of Non-Vector Aphid Feeding Behavior On Virus-Infected Host PlantDocument11 pagesModification of Non-Vector Aphid Feeding Behavior On Virus-Infected Host PlantHayuno SakuraNo ratings yet
- Broad-Spectrum Antimicrobial Activity of Secondary Metabolites ProducedDocument10 pagesBroad-Spectrum Antimicrobial Activity of Secondary Metabolites ProducedVechitra LakshmyNo ratings yet
- E4d2 PDFDocument14 pagesE4d2 PDFPS IMMUNOLOGYNo ratings yet
- Microbial Plant Pathogens: Detection and Management in Seeds and PropagulesFrom EverandMicrobial Plant Pathogens: Detection and Management in Seeds and PropagulesNo ratings yet
- Competitive KIEDocument14 pagesCompetitive KIEkontiki500No ratings yet
- Thermodynamic Interpretation of A Dihedral Angle in Composite MaterialsDocument6 pagesThermodynamic Interpretation of A Dihedral Angle in Composite Materialskontiki500No ratings yet
- Jugoslovenska Medicinsk A BiohemijaDocument77 pagesJugoslovenska Medicinsk A Biohemijakontiki500No ratings yet
- Butscher 2016Document12 pagesButscher 2016kontiki500No ratings yet
- 0350 820X0402093BDocument11 pages0350 820X0402093Bkontiki500No ratings yet
- Jugoslovenska Medicinsk A BiohemijaDocument77 pagesJugoslovenska Medicinsk A Biohemijakontiki500No ratings yet
- Jugoslovenska Medicinsk A BiohemijaDocument77 pagesJugoslovenska Medicinsk A Biohemijakontiki500No ratings yet
- 0367 598X0411479SDocument8 pages0367 598X0411479Skontiki500No ratings yet
- Electrochemical Behaviour of The Catalyst With Kaolinite-Bentonite Substrate in WaterDocument12 pagesElectrochemical Behaviour of The Catalyst With Kaolinite-Bentonite Substrate in Waterkontiki500No ratings yet
- DSS SDocument6 pagesDSS Skontiki500No ratings yet
- 0367 598X0411479SDocument8 pages0367 598X0411479Skontiki500No ratings yet
- 1451 93720502079MDocument6 pages1451 93720502079Mkontiki500No ratings yet
- Serum Lipids in Hypothyroid Children: Effect of Disease and Levothyroxine Replacement TherapyDocument8 pagesSerum Lipids in Hypothyroid Children: Effect of Disease and Levothyroxine Replacement Therapykontiki500No ratings yet
- Biochemical Alterations in Patients With Acute LeukaemiaDocument4 pagesBiochemical Alterations in Patients With Acute Leukaemiakontiki500No ratings yet
- Kinetic Determination of Ultramicro Amounts of Thiocyanate: Facta UniversitatisDocument5 pagesKinetic Determination of Ultramicro Amounts of Thiocyanate: Facta Universitatiskontiki500No ratings yet
- 0367 598X0412548CDocument4 pages0367 598X0412548Ckontiki500No ratings yet
- Experiment of Water Aerosol Estimations of Droplet ParametersDocument12 pagesExperiment of Water Aerosol Estimations of Droplet Parameterskontiki500No ratings yet
- Electrochemical Behaviour of The Catalyst With Kaolinite-Bentonite Substrate in WaterDocument12 pagesElectrochemical Behaviour of The Catalyst With Kaolinite-Bentonite Substrate in Waterkontiki500No ratings yet
- Flavonoids From Flower of Linum Capitatum Kit: Facta UniversitatisDocument5 pagesFlavonoids From Flower of Linum Capitatum Kit: Facta Universitatiskontiki500No ratings yet
- REACTIVITY OF SUBSTITUTED BENZIMIDAZOLE SALTSDocument4 pagesREACTIVITY OF SUBSTITUTED BENZIMIDAZOLE SALTSkontiki500No ratings yet
- Study of Steel Protection with Microalloyed Aluminum CoatingDocument7 pagesStudy of Steel Protection with Microalloyed Aluminum Coatingkontiki500No ratings yet
- Synthesis of New Condensed Coumarin Derivatives: Facta UniversitatisDocument4 pagesSynthesis of New Condensed Coumarin Derivatives: Facta Universitatiskontiki500No ratings yet
- Physical parameters of ventilated filter cigarettesDocument15 pagesPhysical parameters of ventilated filter cigaretteskontiki500No ratings yet
- Monitoring of Bacterial Diseases of Button Mushrooms in SerbiaDocument7 pagesMonitoring of Bacterial Diseases of Button Mushrooms in Serbiakontiki500No ratings yet
- Salting-Out Extraction of Catechol AND Hydroquinone From Aqueous Solutions and Urine SamplesDocument7 pagesSalting-Out Extraction of Catechol AND Hydroquinone From Aqueous Solutions and Urine Sampleskontiki500No ratings yet
- Field Efficacy of Different Fungicide Mixtures in Control of Net Blotch On BarleyDocument7 pagesField Efficacy of Different Fungicide Mixtures in Control of Net Blotch On Barleykontiki500No ratings yet
- Global Warming and Greenhouse Gases: Facta UniversitatisDocument11 pagesGlobal Warming and Greenhouse Gases: Facta Universitatiskontiki500No ratings yet
- Incidence of Viruses in Highbush Blueberry in SerbiaDocument6 pagesIncidence of Viruses in Highbush Blueberry in Serbiakontiki500No ratings yet
- Allelopathic Tolerance of Pea Cultivars To Sorghum Halepense L. (Pers.) ExtractsDocument9 pagesAllelopathic Tolerance of Pea Cultivars To Sorghum Halepense L. (Pers.) Extractskontiki500No ratings yet
- Ultrasound Spectrum of Intraductal Papillary NeoplasmsDocument7 pagesUltrasound Spectrum of Intraductal Papillary NeoplasmsdrrahulsshindeNo ratings yet
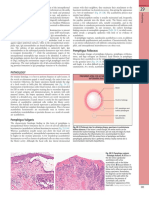
- Pemphigus Foliaceus: Fig. 29.13BDocument1 pagePemphigus Foliaceus: Fig. 29.13BCristian QuitoNo ratings yet
- Interpersonal Relationships Professional Communication Skills For Nurses 6Th Edition Arnold Test Bank Full Chapter PDFDocument29 pagesInterpersonal Relationships Professional Communication Skills For Nurses 6Th Edition Arnold Test Bank Full Chapter PDFhungden8pne100% (11)
- Causes of PtosisDocument3 pagesCauses of PtosisHenry Jonathan BadongenNo ratings yet
- Cardiac Marker Sem 5Document42 pagesCardiac Marker Sem 5novi pujiNo ratings yet
- Surgical SeiveDocument28 pagesSurgical SeiveSumeet PratapNo ratings yet
- Republic of The Philippines Department of Education Region V - BicolDocument2 pagesRepublic of The Philippines Department of Education Region V - BicolKevin Viernes GaraisNo ratings yet
- Kounis Syndrome A Pediatric PerspectiveDocument10 pagesKounis Syndrome A Pediatric PerspectiveAna Belén Artero CastañoNo ratings yet
- N 509 MaterialsDocument128 pagesN 509 MaterialsgdomagasNo ratings yet
- Running Head: QSEN 1Document11 pagesRunning Head: QSEN 1Mariam AbedNo ratings yet
- Usb Powered Led Strip: User GuideDocument5 pagesUsb Powered Led Strip: User Guidespiritos123456789No ratings yet
- Module 2 - 1. Research Overview TranscriptDocument13 pagesModule 2 - 1. Research Overview TranscriptpatrickfvlboltNo ratings yet
- NCP Activity NavidasDocument2 pagesNCP Activity NavidasFran LanNo ratings yet
- Uts Advocacy PaperDocument11 pagesUts Advocacy PaperPlu AldiniNo ratings yet
- NCP For Older Adults With Sleep DisturbanceDocument17 pagesNCP For Older Adults With Sleep DisturbanceAlienda Puspita PutriNo ratings yet
- الميكروبيولوجى والتحكم فى العدوىDocument115 pagesالميكروبيولوجى والتحكم فى العدوىmahmoud salahNo ratings yet
- 1167 1864 1 SMDocument5 pages1167 1864 1 SMMujahid FajrulNo ratings yet
- Genetic Disorders: Chromosomal Abnormalities and Down SyndromeDocument27 pagesGenetic Disorders: Chromosomal Abnormalities and Down SyndromeMerwan KemalNo ratings yet
- Week 3 (20!09!2021) Compe Law ProvisionsDocument18 pagesWeek 3 (20!09!2021) Compe Law ProvisionsNeil FrangilimanNo ratings yet
- KBA Dyslipidemia 2023 Oct 31 TueDocument36 pagesKBA Dyslipidemia 2023 Oct 31 TueKamran B AliNo ratings yet
- Bible Aromatherapy 2022Document39 pagesBible Aromatherapy 2022johanjacobvelosagomezNo ratings yet
- Material Safety Data Sheet - NITRIC ACID PDFDocument9 pagesMaterial Safety Data Sheet - NITRIC ACID PDFJunaid AhmadNo ratings yet
- Pharma Laboratory ExperimentsDocument34 pagesPharma Laboratory Experimentsapi-3748748100% (4)
- Misconceptions About The Use of Genetic Tests in PopulationsDocument4 pagesMisconceptions About The Use of Genetic Tests in PopulationsAneeshNo ratings yet