You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (843)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (346)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- I, Coronavirus. Mother. Monster. Activist. by Bayo AkomolafeDocument64 pagesI, Coronavirus. Mother. Monster. Activist. by Bayo AkomolafeBayo Akomolafe100% (25)
- Essay About COVID 19Document1 pageEssay About COVID 19Diqie Althaf100% (1)
- ENGLISH 7 Quarter 4 Lesson 1Document6 pagesENGLISH 7 Quarter 4 Lesson 1Alleli Faith Leyritana100% (1)
- Population and Sample Practice 1Document3 pagesPopulation and Sample Practice 1Dana_niaNo ratings yet
- Unesco - Eolss Sample Chapters: Statistical Parameter EstimationDocument9 pagesUnesco - Eolss Sample Chapters: Statistical Parameter EstimationDana_niaNo ratings yet
- Introduction To Statistics. An Overview of StatisticsDocument11 pagesIntroduction To Statistics. An Overview of StatisticsDana_niaNo ratings yet
- African Journal of Ecology, Vol 54, Núm 1Document4 pagesAfrican Journal of Ecology, Vol 54, Núm 1Dana_niaNo ratings yet
- Purposes of Data Analysis Parameters and StatisticsDocument20 pagesPurposes of Data Analysis Parameters and StatisticsDana_niaNo ratings yet
- Chapter 15. Nonreligion, Atheism, & Mental HealthDocument24 pagesChapter 15. Nonreligion, Atheism, & Mental HealthDana_niaNo ratings yet
- L1 Belar, C. (2008) (Inglés)Document5 pagesL1 Belar, C. (2008) (Inglés)Dana_niaNo ratings yet
- MacGregor ChristianityOverviewofEarly2016Document7 pagesMacGregor ChristianityOverviewofEarly2016Dana_niaNo ratings yet
- Religions and Politics PDFDocument5 pagesReligions and Politics PDFDana_niaNo ratings yet
- Suicide Silence: The Result Is A Record That's Simultaneously Raw and Intimate, Brutal and PowerfulDocument1 pageSuicide Silence: The Result Is A Record That's Simultaneously Raw and Intimate, Brutal and PowerfulDana_niaNo ratings yet
- Community Based Surveillance Training ManualDocument48 pagesCommunity Based Surveillance Training ManualDana_niaNo ratings yet
- Memory. APA PDFDocument64 pagesMemory. APA PDFDana_niaNo ratings yet
- The Historical Roots of The Pro-Life MovementDocument36 pagesThe Historical Roots of The Pro-Life MovementDana_niaNo ratings yet
- The Long Struggle For The LawDocument6 pagesThe Long Struggle For The LawDana_niaNo ratings yet
- The World's Abortion LawDocument2 pagesThe World's Abortion LawDana_niaNo ratings yet
- COVID-19 Report by Nidhi MithiyaDocument17 pagesCOVID-19 Report by Nidhi MithiyaPooja MithiyaNo ratings yet
- Teks Hortatory CoronaDocument2 pagesTeks Hortatory CoronaNajwa HaifaNo ratings yet
- Coronavirus PDFDocument123 pagesCoronavirus PDFPerla MonroyNo ratings yet
- Con Dential Documents Reveal Moderna Sent mRNA Coronavirus Vaccine Candidate To University Researchers Weeks Before Emergence of Covid-19Document15 pagesCon Dential Documents Reveal Moderna Sent mRNA Coronavirus Vaccine Candidate To University Researchers Weeks Before Emergence of Covid-19hansley cookNo ratings yet
- Social and Preventive Pharmacy BP802 T Semester 8 Unit 2 Imp Question and Answer Unit 2Document23 pagesSocial and Preventive Pharmacy BP802 T Semester 8 Unit 2 Imp Question and Answer Unit 2himmatraojadhao69No ratings yet
- Biology of Sars-Cov-2Document4 pagesBiology of Sars-Cov-2J. L.No ratings yet
- Soal B InggrisDocument20 pagesSoal B Inggrisadnan rifqyNo ratings yet
- Breakingnews & ReadtheoryDocument25 pagesBreakingnews & Readtheoryhanky purnamaNo ratings yet
- Draft List of MicroorganismsDocument36 pagesDraft List of MicroorganismsAngadNo ratings yet
- COVID 19 An Emerging Rapidly Evolving SiDocument8 pagesCOVID 19 An Emerging Rapidly Evolving SigygyNo ratings yet
- Is This The Man Behind The Global Coronavirus PandemicDocument6 pagesIs This The Man Behind The Global Coronavirus PandemicMonte CarloNo ratings yet
- 2020 ImmuneEpitopeMapoftheReportedProteinDocument20 pages2020 ImmuneEpitopeMapoftheReportedProteinvivitri.dewiNo ratings yet
- 'Chapter 1 The Problem and Its BackgroundDocument5 pages'Chapter 1 The Problem and Its BackgroundEinnee Joy CulingNo ratings yet
- Covid ReportDocument1 pageCovid ReportTV UNITNo ratings yet
- Coronavirus Fun Activities Games - 122497Document5 pagesCoronavirus Fun Activities Games - 122497Eve NgewNo ratings yet
- List N Disinfectant Results Table - ExploreDocument72 pagesList N Disinfectant Results Table - ExploreBilal AbbasNo ratings yet
- CONTINGENCY PLAN FOR COVID 19 - A$ SizeDocument15 pagesCONTINGENCY PLAN FOR COVID 19 - A$ Sizemaribel bathanNo ratings yet
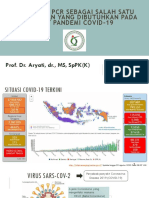
- Multiplex PCR Sebagai Salah Satu Pemeriksaan Yang Dibutuhkan Pada Saat Pandemi Covid-19Document64 pagesMultiplex PCR Sebagai Salah Satu Pemeriksaan Yang Dibutuhkan Pada Saat Pandemi Covid-19Hasna Mirda AmazanNo ratings yet
- 28THFEB2020 - Final - National 2019-nCoV CPDocument34 pages28THFEB2020 - Final - National 2019-nCoV CPMigori Art DataNo ratings yet
- MERS CoVDocument25 pagesMERS CoVJan NerizNo ratings yet
- Covid 19Document59 pagesCovid 19Muhammad SubhanNo ratings yet
- A Study of Plithogenic Graphs: Applications in Spreading Coronavirus Disease (COVID-19) GloballyDocument21 pagesA Study of Plithogenic Graphs: Applications in Spreading Coronavirus Disease (COVID-19) GloballyScience DirectNo ratings yet
- CoViTris2020 and ChloViD2020 A Striking PDFDocument16 pagesCoViTris2020 and ChloViD2020 A Striking PDFgygyNo ratings yet
- Covid 19 PDFDocument117 pagesCovid 19 PDFvicky anandNo ratings yet
- BIBOSO, Christine Marie BILLONES, Beverly Ann BSN4-CDocument30 pagesBIBOSO, Christine Marie BILLONES, Beverly Ann BSN4-CPrecious La-anan BibosoNo ratings yet
- SACS Back To School 2020Document37 pagesSACS Back To School 2020Saige DriverNo ratings yet
- Journal Covid-19Document9 pagesJournal Covid-19NURAIN HANIS BINTI ARIFFNo ratings yet