You might also like
- Bronchial Thermoplasty and Guided Bronchoscopy Part OneDocument25 pagesBronchial Thermoplasty and Guided Bronchoscopy Part OneFistiNo ratings yet
- Jurnal Fungi InhaledDocument13 pagesJurnal Fungi InhaledFistiNo ratings yet
- 10.1016@S1470 20451930318 3Document10 pages10.1016@S1470 20451930318 3FistiNo ratings yet
- Pharmacy Professionals ' Experiences and Perceptions of Providing NHS Patient Medicines Helpline Services: A Qualitative StudyDocument11 pagesPharmacy Professionals ' Experiences and Perceptions of Providing NHS Patient Medicines Helpline Services: A Qualitative StudyFistiNo ratings yet
- Alsaraf 2019 IOP Conf. Ser. Mater. Sci. Eng. 571 012082Document11 pagesAlsaraf 2019 IOP Conf. Ser. Mater. Sci. Eng. 571 012082FistiNo ratings yet
- Characteristics and Outcomes of Acute Kidney Injury in Hospitalized COVID-19 Patients: A Multicenter Study by The Turkish Society of NephrologyDocument25 pagesCharacteristics and Outcomes of Acute Kidney Injury in Hospitalized COVID-19 Patients: A Multicenter Study by The Turkish Society of NephrologyFistiNo ratings yet
- Biopharmaceutics Aspect of Drug Development: Prof. HENNY LUCIDA, PH.D, Apt Andalas UniversityDocument19 pagesBiopharmaceutics Aspect of Drug Development: Prof. HENNY LUCIDA, PH.D, Apt Andalas UniversityFistiNo ratings yet
- Pediatric Parenteral Nutrition Kelly Kopec PharmDocument8 pagesPediatric Parenteral Nutrition Kelly Kopec PharmFistiNo ratings yet
- Patofisiologi: Apt. Najmiatul Fitria, M.Farm., PH.DDocument4 pagesPatofisiologi: Apt. Najmiatul Fitria, M.Farm., PH.DFistiNo ratings yet
- Nilai-Nilai Edukatif Dalam Lirik Nyanyian Onang-Onang Pada Acara Pernikahan Suku Batak Angkola Kabupaten Tapanuli Selatan Provinsi Sumatera UtaraDocument15 pagesNilai-Nilai Edukatif Dalam Lirik Nyanyian Onang-Onang Pada Acara Pernikahan Suku Batak Angkola Kabupaten Tapanuli Selatan Provinsi Sumatera UtaraFistiNo ratings yet
- Ergot Alkaloid BiosynthesisDocument15 pagesErgot Alkaloid BiosynthesisFistiNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- PATOLOGIDocument361 pagesPATOLOGIfe100% (1)
- Topnotch Surgery Supplement HandoutDocument85 pagesTopnotch Surgery Supplement HandoutSarah Michelle NiduaNo ratings yet
- Risk of Opportunistic Infections Associated With Long Term GlucocorticoidDocument20 pagesRisk of Opportunistic Infections Associated With Long Term GlucocorticoidRafael CamachoNo ratings yet
- Impact of Gut Microbiome On Skin Health Gut Skin Axis Observed Through The Lenses of Therapeutics and Skin DiseasesDocument30 pagesImpact of Gut Microbiome On Skin Health Gut Skin Axis Observed Through The Lenses of Therapeutics and Skin DiseasesLudmila Lopes Silva FigueredoNo ratings yet
- Unit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsDocument85 pagesUnit Exam #1: Blood Vessels: B.) Advanced Glycation End ProductsCherry RahimaNo ratings yet
- PhonophoresisDocument22 pagesPhonophoresisnithin nair88% (8)
- Igg4-Related Disease: A Reminder For Practicing PathologistsDocument8 pagesIgg4-Related Disease: A Reminder For Practicing PathologistsMariela Judith UCNo ratings yet
- ODY Efenses: Ap Biology Name Animals Form & Function Activity #4 Date HourDocument5 pagesODY Efenses: Ap Biology Name Animals Form & Function Activity #4 Date Hourancientblackdragon0% (1)
- TOPNOTCH Patho Supplement Handout For Sept 2018 UPDATED May 2018Document25 pagesTOPNOTCH Patho Supplement Handout For Sept 2018 UPDATED May 2018Waiwit KritayakiranaNo ratings yet
- (03241750 - Acta Medica Bulgarica) T Helper Cells in The Immunopathogenesis of Systemic Sclerosis - Current TrendsDocument7 pages(03241750 - Acta Medica Bulgarica) T Helper Cells in The Immunopathogenesis of Systemic Sclerosis - Current TrendsTeodorNo ratings yet
- Cellulite: Etiology, Classification, Pathology, and TreatmentDocument8 pagesCellulite: Etiology, Classification, Pathology, and TreatmentDianaNo ratings yet
- Medical Surgical Nursing - RespiratoryDocument15 pagesMedical Surgical Nursing - RespiratoryChristian Esteves75% (4)
- Gingival EnlargementsDocument34 pagesGingival EnlargementsRohan ShahNo ratings yet
- Taco TraliDocument10 pagesTaco TralimanuelNo ratings yet
- Dry EyeDocument126 pagesDry EyeAnonymous PYgnwOWK100% (4)
- Review Article Andrographis Paniculata (Burm. F.) Wall. Ex NeesDocument28 pagesReview Article Andrographis Paniculata (Burm. F.) Wall. Ex NeesKhalijahNo ratings yet
- Stenotrophomona Maltophilia y Bulkorelia Cepacia. Mnadell 2009Document8 pagesStenotrophomona Maltophilia y Bulkorelia Cepacia. Mnadell 2009Laura López Del Castillo LalydelcaNo ratings yet
- Evolution of Nonsteroidal Anti-Inflammatory Drugs (Nsaids) : Cyclooxygenase (Cox) Inhibition and BeyondDocument30 pagesEvolution of Nonsteroidal Anti-Inflammatory Drugs (Nsaids) : Cyclooxygenase (Cox) Inhibition and BeyondnesakusumaNo ratings yet
- Tsifinalpaper Comment CorrectedDocument43 pagesTsifinalpaper Comment CorrectedTsigereda AberaNo ratings yet
- Tao of Forgotten Food Diet - Taoist Herbology 2Document29 pagesTao of Forgotten Food Diet - Taoist Herbology 2ZioAngelNo ratings yet
- Schuessler's Cell SaltsDocument8 pagesSchuessler's Cell SaltsRoshan Jain MNo ratings yet
- Vitamin C-Lipid Metabolites: Uptake and Retention and Effect On Plasma C-Reactive Protein and Oxidized LDL Levels in Healthy VolunteersDocument5 pagesVitamin C-Lipid Metabolites: Uptake and Retention and Effect On Plasma C-Reactive Protein and Oxidized LDL Levels in Healthy VolunteersDhany NurNo ratings yet
- Fphar 11 596099Document17 pagesFphar 11 596099germanNo ratings yet
- Novak Et Al 2011 AllergyDocument10 pagesNovak Et Al 2011 AllergyArturo VeraNo ratings yet
- Maternal Respiratory Sars-Cov-2 Infection in Pregnancy Is Associated With A Robust Inflammatory Response at The Maternal-Fetal InterfaceDocument31 pagesMaternal Respiratory Sars-Cov-2 Infection in Pregnancy Is Associated With A Robust Inflammatory Response at The Maternal-Fetal InterfaceAllan Depieri CataneoNo ratings yet
- Hypersensitivity Word Document-1Document23 pagesHypersensitivity Word Document-1Dan 04No ratings yet
- Psychedelic Drugs in Biomedicine. Review 2017Document14 pagesPsychedelic Drugs in Biomedicine. Review 2017Luis Henrique SalesNo ratings yet
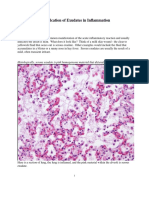
- Identification of Exudates in Inflammation PDFDocument5 pagesIdentification of Exudates in Inflammation PDFlourdes kusumadiNo ratings yet
- PEDIADRUGDocument6 pagesPEDIADRUGPatrice LimNo ratings yet
- Eva-Detko 4-Week ProgramDocument25 pagesEva-Detko 4-Week ProgramVeronika Solntseva100% (3)