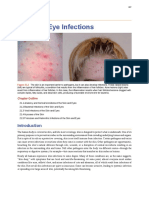
You might also like
- Demodex Mites - Commensals, Parasites or Mutualistic Organisms?Document3 pagesDemodex Mites - Commensals, Parasites or Mutualistic Organisms?SMA Negeri 1 SedayuNo ratings yet
- Demodicosis in Different Age Groups and AlternativDocument23 pagesDemodicosis in Different Age Groups and AlternativJuliana FreitasNo ratings yet
- Reviews: Demodex - An Old Pathogen or A New One?Document4 pagesReviews: Demodex - An Old Pathogen or A New One?Catherine IbtisamahNo ratings yet
- Jurnal Dermatitis Seboroik 3 (Easti)Document8 pagesJurnal Dermatitis Seboroik 3 (Easti)Easti vishiara amdelyNo ratings yet
- Demodex Follicullorum Mimicking Fungal Infection A Case ReportDocument2 pagesDemodex Follicullorum Mimicking Fungal Infection A Case ReportAnonymous V5l8nmcSxb100% (1)
- Facial Dermatosis Associated With DemodexDocument8 pagesFacial Dermatosis Associated With DemodexRoxana SurliuNo ratings yet
- Scalp Seborrheic Dermatitis: What We Know So Far: Skin Appendage DisordersDocument5 pagesScalp Seborrheic Dermatitis: What We Know So Far: Skin Appendage DisordersSergio Revuelta HernándezNo ratings yet
- DandruffDocument13 pagesDandruffYovan PrakosaNo ratings yet
- Fungal InfectionsDocument29 pagesFungal InfectionsKaran MassNo ratings yet
- Microorganisms 09 00543 v2Document19 pagesMicroorganisms 09 00543 v2Pande Agung MahariskiNo ratings yet
- 2nd Half Microbiology For Allied Health StudentsDocument404 pages2nd Half Microbiology For Allied Health StudentsFjornNo ratings yet
- Dermatita SeboreicaDocument4 pagesDermatita SeboreicaFlorian MaghearNo ratings yet
- Dermodex, Estudo CientificoDocument10 pagesDermodex, Estudo CientificoSantinno JrNo ratings yet
- المستند بحث المختبراتDocument3 pagesالمستند بحث المختبراترافت العواضيNo ratings yet
- Perioral DermatitisDocument6 pagesPerioral DermatitisYeni OktaviaNo ratings yet
- SsssDocument5 pagesSsssputeri nashuha100% (1)
- Demo Mite PDF Dermatolgy StudyDocument10 pagesDemo Mite PDF Dermatolgy StudyhLooneygirlNo ratings yet
- Perioral Dermatitis: Acta Dermatovenerologica Croatica: ADC / Hrvatsko Dermatolosko Drustvo February 2008Document6 pagesPerioral Dermatitis: Acta Dermatovenerologica Croatica: ADC / Hrvatsko Dermatolosko Drustvo February 2008Firdani MutiaraNo ratings yet
- DemodexDocument5 pagesDemodexmigas1996No ratings yet
- Perioral Dermatitis PDFDocument6 pagesPerioral Dermatitis PDFanissa fitriNo ratings yet
- Malassezia-Associated Skin Diseases, The Use of Diagnostics and Treatment Saunte2020Document11 pagesMalassezia-Associated Skin Diseases, The Use of Diagnostics and Treatment Saunte2020hannah khumairahNo ratings yet
- Plants Used To Treat Skin Diseases - ReviewDocument9 pagesPlants Used To Treat Skin Diseases - ReviewPsicólogo Clinico clinicoNo ratings yet
- Superficial Mycoses Associated With Diaper DermatitisDocument9 pagesSuperficial Mycoses Associated With Diaper DermatitisIntan FajrinNo ratings yet
- 1 s2.0 S0022202X20311982 Main PDFDocument18 pages1 s2.0 S0022202X20311982 Main PDFKataDullNo ratings yet
- Background: National Rosacea SocietyDocument18 pagesBackground: National Rosacea SocietyPanda OnlineShopNo ratings yet
- Karin Cao Glu 2008Document3 pagesKarin Cao Glu 2008La la landNo ratings yet
- Rosacea 2022Document10 pagesRosacea 2022ucarecufinochiettoNo ratings yet
- DemodexDocument15 pagesDemodexhtandowNo ratings yet
- New Insights Into The Etiopathogenesis of Seborrheic DermatitisDocument5 pagesNew Insights Into The Etiopathogenesis of Seborrheic DermatitisnyomanNo ratings yet
- Mini Review On Dermatomycosis:, C.M. NaziraDocument10 pagesMini Review On Dermatomycosis:, C.M. NaziraKikiNo ratings yet
- Symptomatic Vulvar Demodicosis: A Case Report and Review of The LiteratureDocument4 pagesSymptomatic Vulvar Demodicosis: A Case Report and Review of The LiteratureSilvia Yin CM100% (1)
- Canine Demodicosis - TPIDocument3 pagesCanine Demodicosis - TPIDeepak CNo ratings yet
- WilliamsDocument10 pagesWilliamsTrustia RizqandaruNo ratings yet
- Skin Burns Anatomy Snell Pg. 6Document8 pagesSkin Burns Anatomy Snell Pg. 6CiullaeNo ratings yet
- Microbiom Skin VariabilityDocument5 pagesMicrobiom Skin VariabilityDianaNo ratings yet
- Irritant Dermatitis To Staphylinid Beetle in Indian Troops in CongoDocument4 pagesIrritant Dermatitis To Staphylinid Beetle in Indian Troops in CongorositaNo ratings yet
- Tropical Pathology of The Oral MucosaDocument20 pagesTropical Pathology of The Oral MucosaGeorgi GugicevNo ratings yet
- Seborrheic Dermatitis and Dandruff A Comprehensive Review 2015 PDFDocument10 pagesSeborrheic Dermatitis and Dandruff A Comprehensive Review 2015 PDFAnggriyuni Nur Santi100% (1)
- Darmstadt 1994Document15 pagesDarmstadt 1994rismahNo ratings yet
- Effectiveness of Five Antidandruff Cosmetic Formulations Against Planktonic Cells and Biofilms of DermatophytesDocument7 pagesEffectiveness of Five Antidandruff Cosmetic Formulations Against Planktonic Cells and Biofilms of Dermatophytesjeane wattimenaNo ratings yet
- Isolation and Identification of Malassezia GlobosaDocument8 pagesIsolation and Identification of Malassezia GlobosanahidNo ratings yet
- 1 s2.0 S0923181121000840 MainDocument16 pages1 s2.0 S0923181121000840 Mainleon4009No ratings yet
- Scleroderma - Oral Manifestations and Treatment ChallengesDocument5 pagesScleroderma - Oral Manifestations and Treatment ChallengesLujainNo ratings yet
- Challenges in Exploring and Manipulating The Human Skin Microbiome 2021Document14 pagesChallenges in Exploring and Manipulating The Human Skin Microbiome 2021Vera Brok-VolchanskayaNo ratings yet
- Skin Microbiota: Majalah Kedokteran Sriwijaya, Th. 52 Nomor 1, January 2020Document6 pagesSkin Microbiota: Majalah Kedokteran Sriwijaya, Th. 52 Nomor 1, January 2020Rendra SyaniNo ratings yet
- 5-NOVELTIES IN THE MANAGEMENT OF ROSACEA en 370Document6 pages5-NOVELTIES IN THE MANAGEMENT OF ROSACEA en 370Brîndușa PetruțescuNo ratings yet
- Wanat2015 Cutaneous SarcoidosisDocument18 pagesWanat2015 Cutaneous SarcoidosisJohan ArocaNo ratings yet
- Role of The Skin Microbiome in Dermatology: A LiteraturereviewDocument12 pagesRole of The Skin Microbiome in Dermatology: A LiteraturereviewIJAR JOURNALNo ratings yet
- Pique Ro Casals 2017Document5 pagesPique Ro Casals 2017afrisiammyNo ratings yet
- Mycology - Chapter Six Dimorphic Fungi: BLASTOMYCOSIS (Blastomyces Dermatitidis)Document16 pagesMycology - Chapter Six Dimorphic Fungi: BLASTOMYCOSIS (Blastomyces Dermatitidis)ginarsih99_hutamiNo ratings yet
- Staphylococcal-Scalded Skin Syndrome: Evaluation, Diagnosis, and ManagementDocument6 pagesStaphylococcal-Scalded Skin Syndrome: Evaluation, Diagnosis, and ManagementM Daffa Abhista ReviansyahNo ratings yet
- Herbal Plants and Propionibacterium Acnes - An OverviewDocument13 pagesHerbal Plants and Propionibacterium Acnes - An OverviewmskchoudhuriNo ratings yet
- Darier Disease A Fold (Intertriginous) DermatosisDocument5 pagesDarier Disease A Fold (Intertriginous) Dermatosisdaniel ayllonNo ratings yet
- Dandruff PDFDocument8 pagesDandruff PDFArthur ChietraNo ratings yet
- Loa LoaDocument22 pagesLoa LoaphilmonNo ratings yet
- 07 - Balic NDocument11 pages07 - Balic NK HoNo ratings yet
- 610-Article Text-3639-1-10-20220823Document7 pages610-Article Text-3639-1-10-20220823Gaema MiripNo ratings yet
- Study On Pityriasis Versicolor in Patients Referred To Clinics in TabrizDocument5 pagesStudy On Pityriasis Versicolor in Patients Referred To Clinics in TabrizMiranda Yudhi CyubidubiduuNo ratings yet
- Timer Relay ERV-09Document1 pageTimer Relay ERV-09wal idNo ratings yet
- Chapter 5: Sampling Distributions: Solve The ProblemDocument4 pagesChapter 5: Sampling Distributions: Solve The ProblemEunice WongNo ratings yet
- Feature Writing EnglishDocument2 pagesFeature Writing EnglishAldren BababooeyNo ratings yet
- Stress Management PPT FinalDocument7 pagesStress Management PPT FinalAdarsh Meher100% (1)
- Catalogue: Packer SystemDocument56 pagesCatalogue: Packer SystemChinmoyee Sharma100% (1)
- TC AdvisoryDocument1 pageTC AdvisoryJerome DelfinoNo ratings yet
- K&J Magnetics - Demagnetization CurvesDocument4 pagesK&J Magnetics - Demagnetization CurvessubbannachrsNo ratings yet
- Airbus A319-A320-A321 - Quick Reference HandbookDocument113 pagesAirbus A319-A320-A321 - Quick Reference Handbookrafaelldalpra100% (9)
- Celly BoostbkDocument15 pagesCelly BoostbknomikabNo ratings yet
- IsdettaDocument2 pagesIsdettaHa Phuoc HoaNo ratings yet
- Komatsu Technical BrochurDocument7 pagesKomatsu Technical BrochurBenjamin MossoNo ratings yet
- Leta-Leta CaveDocument5 pagesLeta-Leta CaveToniNo ratings yet
- PECI 405 ECPP 7th Sem CivilDocument96 pagesPECI 405 ECPP 7th Sem CivilYasaswi AkkirajuNo ratings yet
- Jib Crane Assembly ManualDocument76 pagesJib Crane Assembly ManualRobert Cumpa100% (1)
- Age ProblemDocument31 pagesAge ProblemKenny CantilaNo ratings yet
- Macleod - 1974 - Lucian's Knowledge of TheophrastusDocument2 pagesMacleod - 1974 - Lucian's Knowledge of TheophrastusSIMONE BLAIRNo ratings yet
- Lcnews227 - Nexera SeriesDocument47 pagesLcnews227 - Nexera SeriesMuhammad RohmanNo ratings yet
- Visedo FPC-2016Document13 pagesVisedo FPC-2016Probonogoya Erawan SastroredjoNo ratings yet
- AnamnezaDocument3 pagesAnamnezaTeodora StevanovicNo ratings yet
- Manual Generador CAT C15 IbaguéDocument6 pagesManual Generador CAT C15 IbaguéAndres VargasNo ratings yet
- Julie RiesenweberDocument14 pagesJulie RiesenweberisidorabilbaoNo ratings yet
- 132kV Substation Feasibility StudyDocument16 pages132kV Substation Feasibility StudyTafadzwa MurwiraNo ratings yet
- NURS1108 Lecture 10 - Nervous System ENHANCEDDocument40 pagesNURS1108 Lecture 10 - Nervous System ENHANCEDJacia’s SpaceshipNo ratings yet
- Column c4 From 3rd FloorDocument1 pageColumn c4 From 3rd Floor1man1bookNo ratings yet
- Texas Instruments FootprintsDocument7 pagesTexas Instruments FootprintsSteve SmithNo ratings yet
- Additive Manufacturing Objective QuestionsDocument7 pagesAdditive Manufacturing Objective Questionsmohammad shaqib100% (4)
- Design A Roller Coaster ProjectDocument4 pagesDesign A Roller Coaster Projectapi-3564628400% (1)
- ONGC Buyout GOI's Entire 51.11% Stake in HPCLDocument4 pagesONGC Buyout GOI's Entire 51.11% Stake in HPCLArpan AroraNo ratings yet
- 100 Yer PM PPM 0605Document40 pages100 Yer PM PPM 0605biplabpal2009No ratings yet
- Brock Planetary Declination SDocument6 pagesBrock Planetary Declination SDositheus Seth100% (2)