You might also like
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Environmental ResearchDocument12 pagesEnvironmental ResearchnpidasNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- How Many Altogether?: Class 1 3.additionDocument2 pagesHow Many Altogether?: Class 1 3.additionnpidasNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- A-Different-Kind-Of-School NotesDocument2 pagesA-Different-Kind-Of-School NotesnpidasNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Addition Worksheet Class 1Document2 pagesAddition Worksheet Class 1npidasNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- A Happy Child: Text Based QuestionsDocument6 pagesA Happy Child: Text Based QuestionsSUBHANo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Potassium Nutrition in Plants and Its Interactions With Other Nutrients in Hydroponic CultureDocument14 pagesPotassium Nutrition in Plants and Its Interactions With Other Nutrients in Hydroponic CulturenpidasNo ratings yet
- Muller's Nobel Lecture On Dose-Response For Ionizing Radiation: Ideology or Science?Document4 pagesMuller's Nobel Lecture On Dose-Response For Ionizing Radiation: Ideology or Science?npidasNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- 1 STD Roll and SlideDocument1 page1 STD Roll and SlidenpidasNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Sustainable Initiative of Using Cyanobacteria As A Liquid Fertilizer For Hydroponic Cultivation: A Waste To Wealth UtilizationDocument32 pagesSustainable Initiative of Using Cyanobacteria As A Liquid Fertilizer For Hydroponic Cultivation: A Waste To Wealth UtilizationnpidasNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Environmental Technology & Innovation: Uday Kumar Banala, Nilamadhab Prasad Indradyumna Das, Subba Rao ToletiDocument17 pagesEnvironmental Technology & Innovation: Uday Kumar Banala, Nilamadhab Prasad Indradyumna Das, Subba Rao ToletinpidasNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
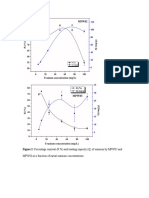
- Figure 2: Percentage Removal (R %) and Loading Capacity (Q) of Uranium by MPW02 and MPW03 As A Function of Initial Uranium ConcentrationsDocument6 pagesFigure 2: Percentage Removal (R %) and Loading Capacity (Q) of Uranium by MPW02 and MPW03 As A Function of Initial Uranium ConcentrationsnpidasNo ratings yet
- 3.5. FT-IR AnalysisDocument5 pages3.5. FT-IR AnalysisnpidasNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- P 2Document5 pagesP 2npidasNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Kinetics of Uranium Sequestration by Two Natural Isolates From Alkaline Uranium Mine WatersDocument5 pagesKinetics of Uranium Sequestration by Two Natural Isolates From Alkaline Uranium Mine WatersnpidasNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Microbial Interaction With U JTEDocument46 pagesMicrobial Interaction With U JTEnpidasNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Thrigmopoeus PsychedelicusDocument7 pagesThrigmopoeus PsychedelicusnpidasNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Environmental Technology & InnovationDocument12 pagesEnvironmental Technology & InnovationnpidasNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- DrosophilaDocument18 pagesDrosophilanpidasNo ratings yet
- Uday JHMPaper 13 JanDocument11 pagesUday JHMPaper 13 JannpidasNo ratings yet
- Detection of Air CovidDocument7 pagesDetection of Air CovidnpidasNo ratings yet
- Safety Measures To Prevent SARS-CoV-2 Laboratory-Acquired Infection - Clinical Lab ManagerDocument9 pagesSafety Measures To Prevent SARS-CoV-2 Laboratory-Acquired Infection - Clinical Lab ManagernpidasNo ratings yet
- A Coordinated Global Research Roadmap:: 2019 Novel CoronavirusDocument36 pagesA Coordinated Global Research Roadmap:: 2019 Novel CoronavirusManuel OropezaNo ratings yet
- Key Clinical Research Priorities For The Pediatric Community During The COVID-19 PandemicDocument3 pagesKey Clinical Research Priorities For The Pediatric Community During The COVID-19 PandemicnpidasNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Immunogenicity of A DNA Vaccine Candidate For COVID-19: ArticleDocument13 pagesImmunogenicity of A DNA Vaccine Candidate For COVID-19: ArticlePropllusNo ratings yet
- Radiation BiologyDocument166 pagesRadiation BiologyAnh Tran100% (4)
- Serum Iga, Igm, and Igg Responses in Covid-19: Cellular & Molecular ImmunologyDocument3 pagesSerum Iga, Igm, and Igg Responses in Covid-19: Cellular & Molecular ImmunologynpidasNo ratings yet
- Radiation BiologyDocument166 pagesRadiation BiologyAnh Tran100% (4)
- 9th Science EMDocument336 pages9th Science EMKalai ArasiNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Wetland Birds of KalpakkamDocument11 pagesWetland Birds of KalpakkamnpidasNo ratings yet
- 2 NdresearchpaperDocument5 pages2 Ndresearchpaperapi-356685046No ratings yet
- IELTS READING TEST 33: Bilingualism Advantages and Changing Health RulesDocument12 pagesIELTS READING TEST 33: Bilingualism Advantages and Changing Health RulesDollar KhokharNo ratings yet
- Directory 18-08-21Document34 pagesDirectory 18-08-21api-309412486100% (1)
- Assignment On HealthDocument13 pagesAssignment On HealthMit MecwanNo ratings yet
- Fate of Manganese Associated With The Inhalation of Welding Fumes: Potential Neurological EffectsDocument7 pagesFate of Manganese Associated With The Inhalation of Welding Fumes: Potential Neurological EffectsAndrea Carolina Salas LalindeNo ratings yet
- Diag AutismoDocument12 pagesDiag AutismoSandra Carranza ZavalaNo ratings yet
- Researchers and by Clinicians For Use With Their PatientsDocument3 pagesResearchers and by Clinicians For Use With Their PatientsAnton Henry MiagaNo ratings yet
- 18.20.04 - BSSL Seafarers Health Self Declaration With COVID 19 Test And... (1) - 3Document3 pages18.20.04 - BSSL Seafarers Health Self Declaration With COVID 19 Test And... (1) - 3Dean IL RE JamesNo ratings yet
- HOPE 1 Module 8Document2 pagesHOPE 1 Module 8Kevin Viernes Garais100% (2)
- How To Dramatically Increase Your Vocal Range PDFDocument25 pagesHow To Dramatically Increase Your Vocal Range PDFjoaosilvaNo ratings yet
- Power Point Consumer Health EducationDocument81 pagesPower Point Consumer Health EducationAlbert Ian CasugaNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- SV The Coronation of King Charles IIIDocument9 pagesSV The Coronation of King Charles IIIMaria GromovaNo ratings yet
- Best Body FitnessDocument3 pagesBest Body FitnessLavinia Chirila-StefanutNo ratings yet
- This Study Determined The Effectiveness of Mnemonics in Improving Memory Among Nursing Students in GCON, BikanerDocument3 pagesThis Study Determined The Effectiveness of Mnemonics in Improving Memory Among Nursing Students in GCON, BikanerNaveen ShrivasNo ratings yet
- Nursing Board Exam Answer Key and Test QuestionsDocument44 pagesNursing Board Exam Answer Key and Test Questionsrem_cen24100% (1)
- Re-Opening Plan For The 2020-2021 School YearDocument18 pagesRe-Opening Plan For The 2020-2021 School YearNews 8 WROCNo ratings yet
- Physical Education 9: (Learner's Material) Quarter 1Document185 pagesPhysical Education 9: (Learner's Material) Quarter 1Lianne Grace De VeraNo ratings yet
- Effectiveness of T-Piece Resuscitator Versus Self-Inflating Bag During Birth Resuscitation in Very Low Birth Weight InfantsDocument7 pagesEffectiveness of T-Piece Resuscitator Versus Self-Inflating Bag During Birth Resuscitation in Very Low Birth Weight InfantsRaja SekarNo ratings yet
- Personal Values, Beliefs and AttitudesDocument10 pagesPersonal Values, Beliefs and Attitudesapi-282066793No ratings yet
- Skilling For LivelihoodDocument19 pagesSkilling For LivelihoodPawanKumarNo ratings yet
- Expected Coding and Decoding Questions For Ibps Po MainsDocument11 pagesExpected Coding and Decoding Questions For Ibps Po Mainsvirat kohli86% (7)
- (EBPH) : Mengembangkan Evidence Based Public Health Hiv Dan Aids Berbasis SurveilansDocument8 pages(EBPH) : Mengembangkan Evidence Based Public Health Hiv Dan Aids Berbasis SurveilansDamiriNo ratings yet
- Ultimate Yogi CalendarDocument1 pageUltimate Yogi CalendarDavidPérez100% (3)
- The Hall Technique 10 Years On: Questions and Answers: GeneralDocument6 pagesThe Hall Technique 10 Years On: Questions and Answers: GeneralamilescoNo ratings yet
- College of Liberal Arts University of LuzonDocument9 pagesCollege of Liberal Arts University of LuzonChristianne Joyse MerreraNo ratings yet
- ORGB 5th Edition Nelson Solutions Manual 1Document44 pagesORGB 5th Edition Nelson Solutions Manual 1shirley100% (52)
- Abstract of ThesisDocument2 pagesAbstract of ThesisdrkartikeytyagiNo ratings yet
- Long-Term Abstinence Following Holotropic Breathwork As Adjunctive Treatment of Substance Use Disorders and Related Psychiatric ComorbidityDocument7 pagesLong-Term Abstinence Following Holotropic Breathwork As Adjunctive Treatment of Substance Use Disorders and Related Psychiatric ComorbiditymghuesoNo ratings yet
- OSHA Complaint FinalDocument3 pagesOSHA Complaint FinalBethanyNo ratings yet
- Sports Day Narrative Report 2022-2023Document6 pagesSports Day Narrative Report 2022-2023Bloom Sarinas100% (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)