You might also like
- Artigo Melasma Happy HourDocument14 pagesArtigo Melasma Happy HourAline Brito Ferreira de CastroNo ratings yet
- Current Concepts in MelasmaDocument5 pagesCurrent Concepts in MelasmaNikhil ShrivastavaNo ratings yet
- Melasma A Prospective Study of Clinico Etiological and Dermoscopic Pattern in MenDocument4 pagesMelasma A Prospective Study of Clinico Etiological and Dermoscopic Pattern in MenInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Melasma UpdateDocument11 pagesMelasma Updateirwan jun100% (1)
- The Association Between Melasma and Postin Ammatory Hyperpigmenta-Tion in Acne PatientsDocument4 pagesThe Association Between Melasma and Postin Ammatory Hyperpigmenta-Tion in Acne PatientsRicoNo ratings yet
- Melasma Men 2012Document8 pagesMelasma Men 2012Luci Ramona Alvarenga BenitezNo ratings yet
- Glacial RXDocument15 pagesGlacial RXcarloshgmedeirosNo ratings yet
- DamevskamelasmasjdvDocument15 pagesDamevskamelasmasjdvlaithuyduong007No ratings yet
- PCMR 12404Document14 pagesPCMR 12404Riefka Ananda ZulfaNo ratings yet
- New Oral and Topical Approaches For The Treatment of MelasmaDocument7 pagesNew Oral and Topical Approaches For The Treatment of MelasmaSong JieunNo ratings yet
- International Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakDocument7 pagesInternational Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakRiefka Ananda ZulfaNo ratings yet
- Melasma Pathogenesis and Influencing FactorsDocument2 pagesMelasma Pathogenesis and Influencing FactorsTatianaNo ratings yet
- Malassezia FolliculitisDocument5 pagesMalassezia Folliculitisshivam18No ratings yet
- Sarkar, RashmiDocument10 pagesSarkar, RashmiXena Poetri Xaverya RenggaNo ratings yet
- Focus Doolan MelasmaDocument7 pagesFocus Doolan MelasmaEnderson MedeirosNo ratings yet
- Jurnal Keperluan PentingDocument4 pagesJurnal Keperluan PentingmhdtaufikkNo ratings yet
- Update PatogenesisDocument22 pagesUpdate PatogenesisUI KipediaNo ratings yet
- 10 1111@jocd 12877Document6 pages10 1111@jocd 12877ohmanhbkNo ratings yet
- Melasma: An Up-to-Date Comprehensive Review: Oluwatobi A. Ogbechie-Godec Nada ElbulukDocument14 pagesMelasma: An Up-to-Date Comprehensive Review: Oluwatobi A. Ogbechie-Godec Nada ElbulukDra Natalia Maria ZuluagaNo ratings yet
- 2021 The Pathogenesis of Melasma and Implications For TreatmentDocument14 pages2021 The Pathogenesis of Melasma and Implications For Treatmentyezafig4001No ratings yet
- 115Document14 pages115Jowi Tsidkenu Pili CruzNo ratings yet
- PoikilodermaDocument5 pagesPoikilodermaDiana NunesNo ratings yet
- PRELIMS - Indd 17 1/18/2018 11:06:42 AMDocument7 pagesPRELIMS - Indd 17 1/18/2018 11:06:42 AMhoang ho LeNo ratings yet
- Diagnostics 11 00794 v2Document11 pagesDiagnostics 11 00794 v2Adn crnNo ratings yet
- A Comparative Study of Efficacy Between Modified Kligmans Formula and Dermaroller With Tranexamic Acid in Treatment of MelasmaDocument8 pagesA Comparative Study of Efficacy Between Modified Kligmans Formula and Dermaroller With Tranexamic Acid in Treatment of MelasmaIJAR JOURNALNo ratings yet
- Tratment Melasma PDFDocument18 pagesTratment Melasma PDFGheavita Chandra DewiNo ratings yet
- 10.11648.j.ijcd.20170102.12 2Document8 pages10.11648.j.ijcd.20170102.12 2Muhammad UmerNo ratings yet
- MelaFind Study Shows High Sensitivity for Detecting MelanomaDocument8 pagesMelaFind Study Shows High Sensitivity for Detecting MelanomadiiinnaliinnnNo ratings yet
- Melasma: Continuing Education ActivityDocument9 pagesMelasma: Continuing Education ActivityEldie RahimNo ratings yet
- Pigment Int EngimaofcolorintineaversicolorDocument5 pagesPigment Int EngimaofcolorintineaversicolorKuldeepSinghBanaNo ratings yet
- An Overview On Melasma 2376 0427 1000216Document18 pagesAn Overview On Melasma 2376 0427 1000216dhilaidrisNo ratings yet
- An Evidence-Based Approach To Pediatric MelanonychiaDocument13 pagesAn Evidence-Based Approach To Pediatric MelanonychiaMarice QCNo ratings yet
- Clinico Morphological Evaluation of Melasma and Other Hyperpigmentary Disorders of SkinDocument25 pagesClinico Morphological Evaluation of Melasma and Other Hyperpigmentary Disorders of SkinBaru Chandrasekhar RaoNo ratings yet
- Journal-Of-Dermatology-Research-And-Therapy-Ijdrt-6-090Document6 pagesJournal-Of-Dermatology-Research-And-Therapy-Ijdrt-6-090seydou.organismNo ratings yet
- Contoh IHK OKDocument7 pagesContoh IHK OKenolabrown11No ratings yet
- Melasma Insights and PerspectivesDocument6 pagesMelasma Insights and PerspectivesTatianaNo ratings yet
- Thyroid Disorders in Patients With MelasmaDocument8 pagesThyroid Disorders in Patients With MelasmaIJAR JOURNALNo ratings yet
- Advances in Malignant Melanoma - Clinical and Research PerspectivesDocument264 pagesAdvances in Malignant Melanoma - Clinical and Research PerspectivesInternational Medical PublisherNo ratings yet
- Neagu 2021Document40 pagesNeagu 2021meenakshi.r.agrawalNo ratings yet
- Hyperpigmentation and Melasma: D Rigopoulos, MD, PHD, S Gregoriou, MD, & A Katsambas, MD, PHDDocument8 pagesHyperpigmentation and Melasma: D Rigopoulos, MD, PHD, S Gregoriou, MD, & A Katsambas, MD, PHDGÜLSÜM GÜNEŞNo ratings yet
- PCR Tugas Diagmol 2Document9 pagesPCR Tugas Diagmol 2Laboratorium RSUDKOTANGNo ratings yet
- Endometrial Gene Expression: An Emerging Paradigm for Reproductive DisordersFrom EverandEndometrial Gene Expression: An Emerging Paradigm for Reproductive DisordersJoanne Kwak-KimNo ratings yet
- MelasmDocument22 pagesMelasmLeonardo Alves F AlmeidaNo ratings yet
- Dlova Et Al-2018-British Journal of DermatologyDocument2 pagesDlova Et Al-2018-British Journal of DermatologyAndrea CordaroNo ratings yet
- Frontal Fbrosing Alopecia and Personal Care Product Use - A Systematic Review and Meta AnalysisDocument19 pagesFrontal Fbrosing Alopecia and Personal Care Product Use - A Systematic Review and Meta AnalysisAna MariaNo ratings yet
- Meds CapeDocument14 pagesMeds CapeMeta SakinaNo ratings yet
- En V58n5a22Document13 pagesEn V58n5a22M Nurshaffriaal RusdiNo ratings yet
- Prevalence and Quality of Life of Pemphigus Patients at Sanglah General Hospital Bali-IndonesiaDocument4 pagesPrevalence and Quality of Life of Pemphigus Patients at Sanglah General Hospital Bali-Indonesiashat bidNo ratings yet
- Skin Ageing: Abstracts / Maturitas 81 (2015) 191-236 233Document1 pageSkin Ageing: Abstracts / Maturitas 81 (2015) 191-236 233Yunita AmiliaNo ratings yet
- Evaluation and Management of Refractory Acne Vulgaris in Adolescent and Adult MenDocument4 pagesEvaluation and Management of Refractory Acne Vulgaris in Adolescent and Adult MenKadir KUCUKNo ratings yet
- A Case Report of A Morphea in A 19 Years Old FemaleDocument2 pagesA Case Report of A Morphea in A 19 Years Old FemaleInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Wagner Et Al., 2002 (Comparing Quantitative Measures)Document6 pagesWagner Et Al., 2002 (Comparing Quantitative Measures)Aurelia SuryaniNo ratings yet
- Dr. Tandiono Hermanda, SP - KKDocument6 pagesDr. Tandiono Hermanda, SP - KKAchmad Fahri BaharsyahNo ratings yet
- Cosmetics 10 00025 v2 - RemovedDocument2 pagesCosmetics 10 00025 v2 - Removedmeisya 102018090No ratings yet
- Lifetime prevalence and characteristics of photodermatoses in EuropeDocument1 pageLifetime prevalence and characteristics of photodermatoses in EuropekelbmutsNo ratings yet
- Cme Reviewarticle: The Role of Genes in The Development of Mullerian AnomaliesDocument9 pagesCme Reviewarticle: The Role of Genes in The Development of Mullerian Anomaliesapi-54418266No ratings yet
- Cutaneous MelanomaDocument15 pagesCutaneous MelanomadaltondittzNo ratings yet
- AJGP 12 2021 Focus Doolan Melasma WEBDocument6 pagesAJGP 12 2021 Focus Doolan Melasma WEBضحى الغزيNo ratings yet
- Laino 2018Document9 pagesLaino 2018Henny WijayaNo ratings yet
- Genetics of Pre-Eclampsia and The Weight of Babies at Birth - Clinical and Genetic Studies in Sri LankaDocument3 pagesGenetics of Pre-Eclampsia and The Weight of Babies at Birth - Clinical and Genetic Studies in Sri LankaMuhammad FaisalNo ratings yet
- Acs Chemrestox 0c00461Document9 pagesAcs Chemrestox 0c00461FABIANA PASSAMANINo ratings yet
- Role of nutrients in immunityDocument68 pagesRole of nutrients in immunityNatta ApucaranaNo ratings yet
- 21 Artigo Magnesio e Respiracao Celular CompletoDocument8 pages21 Artigo Magnesio e Respiracao Celular CompletoNatta ApucaranaNo ratings yet
- 34 Vit D Antiox I CompletoDocument7 pages34 Vit D Antiox I CompletoNatta ApucaranaNo ratings yet
- Research Article: Relationship Between Vitamin D Status and Striae Distensae: A Case-Referent StudyDocument6 pagesResearch Article: Relationship Between Vitamin D Status and Striae Distensae: A Case-Referent StudyNatta ApucaranaNo ratings yet
- 31 Vit D e Calcio Reg Da Cicatrizacao Ferida CompletoDocument7 pages31 Vit D e Calcio Reg Da Cicatrizacao Ferida CompletoNatta ApucaranaNo ratings yet
- MELASMA: Etiology, Clinical Features, Differential Diagnoses and Treatment OptionsDocument20 pagesMELASMA: Etiology, Clinical Features, Differential Diagnoses and Treatment OptionsAndi Firman MubarakNo ratings yet
- Gerda Pelayo - MELASMA TREATMENT SETDocument1 pageGerda Pelayo - MELASMA TREATMENT SETVictoria Carl CortinaNo ratings yet
- IPL - Skin Rejuvenation - PROTOCOL PDFDocument45 pagesIPL - Skin Rejuvenation - PROTOCOL PDFmochkurniawan83% (6)
- WefdfafasDocument3 pagesWefdfafasPatrick AllenNo ratings yet
- Skin Color Types and Indian Skin CharacteristicsCh-4Document16 pagesSkin Color Types and Indian Skin CharacteristicsCh-4Nilavan MuthuNo ratings yet
- HIPERPIGMENTASI TREATMENT OPTIONSDocument14 pagesHIPERPIGMENTASI TREATMENT OPTIONSpuskesmas tanjungharjoNo ratings yet
- Tranexamic Acid For Melasma TreatmentDocument6 pagesTranexamic Acid For Melasma TreatmentBeatriz CristinaNo ratings yet
- Skin Problems Multiple Choice QuestionsDocument9 pagesSkin Problems Multiple Choice QuestionsAliNo ratings yet
- A Stepwise Approach To Melasma Management and TreatmentDocument8 pagesA Stepwise Approach To Melasma Management and TreatmentMeta SakinaNo ratings yet
- Update On Melasma-Part II: TreatmentDocument24 pagesUpdate On Melasma-Part II: TreatmentTrần Hồng MyNo ratings yet
- (PDF) Future Therapies in Melasma: What Lies Ahead?Document16 pages(PDF) Future Therapies in Melasma: What Lies Ahead?Riefka Ananda ZulfaNo ratings yet
- Dermoscopy Serves Dual Role in Melasma ManagementDocument22 pagesDermoscopy Serves Dual Role in Melasma ManagementMeta SakinaNo ratings yet
- 2017 - Efficacy of Cysteamine Cream in The Treatment of Epidermal Melasma, Evaluating by Dermacatch As A New Measurement Method. A Randomized Double Blind Placebo Controlled Study.Document9 pages2017 - Efficacy of Cysteamine Cream in The Treatment of Epidermal Melasma, Evaluating by Dermacatch As A New Measurement Method. A Randomized Double Blind Placebo Controlled Study.Adriana GomesNo ratings yet
- Topical and Oral Melatonin for Melasma ManagementDocument15 pagesTopical and Oral Melatonin for Melasma ManagementdeswitrigintaNo ratings yet
- Pigmented Contact Dermatitis 2376 0427 1000214Document4 pagesPigmented Contact Dermatitis 2376 0427 1000214iqbal dsNo ratings yet
- Exd 13915Document10 pagesExd 13915dominiqueNo ratings yet
- v3 Avaliacao Da Eficacia Do Licopeno Beta Caroteno e Lactobacillus Johnsonii No Tratamento de Manutencao Do Melasma Durante o Verao Um Estudo ComparativoDocument5 pagesv3 Avaliacao Da Eficacia Do Licopeno Beta Caroteno e Lactobacillus Johnsonii No Tratamento de Manutencao Do Melasma Durante o Verao Um Estudo Comparativohendro wNo ratings yet
- Get Rid of Dark Spots - The Power of Homeopathy For Pigmentation - Schwabe IndiaDocument11 pagesGet Rid of Dark Spots - The Power of Homeopathy For Pigmentation - Schwabe Indianadee.ak2023No ratings yet
- PigmentationDocument4 pagesPigmentationDavid DijakNo ratings yet
- The Effectiveness of Combination Serum of Tranexamic Acid, Galactomyces Ferment Filtrate, Niacinamide and Alpha Arbutin in Enhancing Skin BrightnessDocument5 pagesThe Effectiveness of Combination Serum of Tranexamic Acid, Galactomyces Ferment Filtrate, Niacinamide and Alpha Arbutin in Enhancing Skin BrightnessFachrizal AuliaNo ratings yet
- DERM NotesDocument19 pagesDERM NoteszakyNo ratings yet
- Federico Omolon - Melasma Treatment SetDocument2 pagesFederico Omolon - Melasma Treatment SetVictoria Carl CortinaNo ratings yet
- The Versatility of Azelaic Acid in DermatologyDocument32 pagesThe Versatility of Azelaic Acid in DermatologyGourav SainNo ratings yet
- Exogenous Ochronosis-Successful Outcome After Treatment QS NdYAG LaserDocument6 pagesExogenous Ochronosis-Successful Outcome After Treatment QS NdYAG LaseradeNo ratings yet
- Medicine: Efficacy of Functional Microarray of Microneedles Combined With Topical Tranexamic Acid For MelasmaDocument7 pagesMedicine: Efficacy of Functional Microarray of Microneedles Combined With Topical Tranexamic Acid For MelasmaPin Han NaNo ratings yet
- International Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakDocument7 pagesInternational Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakRiefka Ananda ZulfaNo ratings yet
- Skin Pigmentation + Hari DisorderDocument113 pagesSkin Pigmentation + Hari DisorderAfiqah So JasmiNo ratings yet
- Intervention For Melasma PDFDocument87 pagesIntervention For Melasma PDFLong HoàngNo ratings yet
- Tranexamic Acid in Melasma A Review 2376 0427 1000228Document4 pagesTranexamic Acid in Melasma A Review 2376 0427 1000228Ayu WikanNo ratings yet
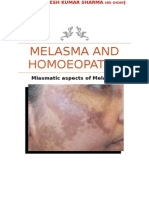
- Melasma and HomoeopathyDocument14 pagesMelasma and HomoeopathyDr. Rajneesh Kumar Sharma MD HomNo ratings yet