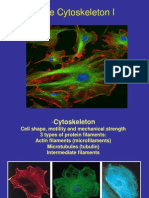
You might also like
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Cytoskeleton: Jetro P. MagnayonDocument66 pagesCytoskeleton: Jetro P. MagnayonChristian Santiago100% (1)
- Cell Bio Final ReviewDocument13 pagesCell Bio Final ReviewFekete LiliNo ratings yet
- 05lect Adhesion Summary2017Document5 pages05lect Adhesion Summary2017Dan ChoiNo ratings yet
- 14 The CytoskeletonDocument7 pages14 The Cytoskeletonsatriamanullang880No ratings yet
- 02lect Cell1 Summary2017Document4 pages02lect Cell1 Summary2017Dan ChoiNo ratings yet
- 11.sitoskeleton Dan Pergerakan Sel-Dr - IsraDocument40 pages11.sitoskeleton Dan Pergerakan Sel-Dr - IsraTa RaNo ratings yet
- Cytoskeletal NetworkDocument51 pagesCytoskeletal Networkconan128No ratings yet
- BY2201 EmmaC CytoskeletonIandIIDocument62 pagesBY2201 EmmaC CytoskeletonIandIIRebecca ReillyNo ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- Cy To SkeletonDocument17 pagesCy To SkeletonKitkat AlorroNo ratings yet
- Cytoskeleton and Cell MotilityDocument47 pagesCytoskeleton and Cell MotilityDyang AprilianiNo ratings yet
- The CytoskeletonDocument63 pagesThe CytoskeletonAmara TargaryenNo ratings yet
- Biologi Sel Idk 1Document30 pagesBiologi Sel Idk 1ImanNo ratings yet
- Extracellular Matrix and CytoskeletonDocument23 pagesExtracellular Matrix and CytoskeletonjoyyeesufNo ratings yet
- Cytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDocument11 pagesCytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDario DiosNo ratings yet
- Lesson 2: Molecular Organization of CellsDocument12 pagesLesson 2: Molecular Organization of CellsremshopNo ratings yet
- The Cytoskeleton: Cell Biology 1st SemesterDocument22 pagesThe Cytoskeleton: Cell Biology 1st SemesterJason CenaNo ratings yet
- Biology ObjectivesDocument12 pagesBiology ObjectivesNicholas OwNo ratings yet
- Biochem AssignmentDocument2 pagesBiochem AssignmentAngel GoNo ratings yet
- CHAP 2. CSH TẾ BÀO - 232Document100 pagesCHAP 2. CSH TẾ BÀO - 232Nguyễn Anh QuangNo ratings yet
- Full Download Human Anatomy 8th Edition Martini Solutions ManualDocument36 pagesFull Download Human Anatomy 8th Edition Martini Solutions Manualberthasauflyvski1j100% (29)
- Dwnload Full Human Anatomy 8th Edition Martini Solutions Manual PDFDocument36 pagesDwnload Full Human Anatomy 8th Edition Martini Solutions Manual PDFcapsicum.imprison0fwm100% (7)
- Prepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemDocument37 pagesPrepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemleartaNo ratings yet
- Lecture 20 - Cell Organization & MotilityDocument3 pagesLecture 20 - Cell Organization & MotilityDana20SNo ratings yet
- Cell Biology Questions and AnswersDocument14 pagesCell Biology Questions and Answersapi-19916399No ratings yet
- PRQ7Document2 pagesPRQ7glody mbokoNo ratings yet
- Cell Structure Mitosis and Meiosis 2007 For UPLOADDocument44 pagesCell Structure Mitosis and Meiosis 2007 For UPLOADJason RobinsonNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- Lecture 6 - Cytoskeleton STDocument18 pagesLecture 6 - Cytoskeleton STminhph.23bi14469No ratings yet
- Molecular Bioeng-2: Mrs Daramola BellstechDocument22 pagesMolecular Bioeng-2: Mrs Daramola BellstechSamuel EkpoNo ratings yet
- CytoplasmDocument13 pagesCytoplasmTanveerNo ratings yet
- Bookmark This Page: Cadherins Mucin-Like Cams IntegrinsDocument38 pagesBookmark This Page: Cadherins Mucin-Like Cams IntegrinsAndreea SpiridonNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- Through Which Ions, Sugars, Amino Acids, and Other Small Molecules May PassDocument2 pagesThrough Which Ions, Sugars, Amino Acids, and Other Small Molecules May Passan1216073No ratings yet
- Clinical Case #1 Botulism 1. Discuss The Functional Unit of Muscle and Its MyofibrilsDocument3 pagesClinical Case #1 Botulism 1. Discuss The Functional Unit of Muscle and Its MyofibrilsArdette De JesusNo ratings yet
- Emt318 Cytology Note3 2021Document10 pagesEmt318 Cytology Note3 2021Ehigie promiseNo ratings yet
- 2cytology Cytoskel Receptor Transp (3) 1Document43 pages2cytology Cytoskel Receptor Transp (3) 1m.mansoor3377No ratings yet
- 04 Structure To FunctionDocument5 pages04 Structure To Function897291868100% (1)
- The Cell - Cytosol and Cytoskeleton - LecturioDocument5 pagesThe Cell - Cytosol and Cytoskeleton - LecturiosylviemwaluleNo ratings yet
- Lecture2 110920064308 Phpapp01Document44 pagesLecture2 110920064308 Phpapp01John Louis PulidoNo ratings yet
- Human Anatomy 8th Edition Martini Solutions ManualDocument26 pagesHuman Anatomy 8th Edition Martini Solutions ManualLeslieBuchanannjfy100% (57)
- SJDFSJHDocument3 pagesSJDFSJHapi-296833859No ratings yet
- 4.1 CytoskeletonDocument28 pages4.1 Cytoskeletonedabzan234No ratings yet
- Chapter 16 SummaryDocument11 pagesChapter 16 SummaryCharlotteNo ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- Amino Acids and ProteinsDocument2 pagesAmino Acids and ProteinsyejiNo ratings yet
- Cell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Document26 pagesCell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Rajni KumariNo ratings yet
- 06lect ECM Summary2017Document5 pages06lect ECM Summary2017Dan ChoiNo ratings yet
- 3 - Histology Lecture - Structure of Non Membranous Cell OrganellesDocument42 pages3 - Histology Lecture - Structure of Non Membranous Cell OrganellesAMIRA HELAYELNo ratings yet
- February 27, 2012 - Chapter 7: Inside The CellDocument4 pagesFebruary 27, 2012 - Chapter 7: Inside The CellVincent Troy SantosNo ratings yet
- NUO011Document38 pagesNUO011Hello HiNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- 2016 10 18 The Extracellular Matrix (ECM)Document45 pages2016 10 18 The Extracellular Matrix (ECM)sennaavia12100% (1)
- Topic: Cytokinesis: Etymology and PronunciationDocument6 pagesTopic: Cytokinesis: Etymology and PronunciationVindhya ShankerNo ratings yet
- Basic and Applied Cell Physiology: Reference BooksDocument28 pagesBasic and Applied Cell Physiology: Reference Booksjamila ilNo ratings yet
- BIOL 202 Lab 2 Animal Cells and Tissues: Cell StructureDocument15 pagesBIOL 202 Lab 2 Animal Cells and Tissues: Cell StructureKate Anne Montallana RamosNo ratings yet
- Cellular Nanomechanics 2Document27 pagesCellular Nanomechanics 2Ahmed MotasimNo ratings yet
- Molecular Cell Biology BS101Document35 pagesMolecular Cell Biology BS101omgwtfbbq170070No ratings yet
- Cell Cycle NotesDocument12 pagesCell Cycle NotesMhariane MabborangNo ratings yet
- 11lect Muscle2 Summary2017Document4 pages11lect Muscle2 Summary2017Dan ChoiNo ratings yet
- 13lect Nerve2 Summary2017Document4 pages13lect Nerve2 Summary2017Dan ChoiNo ratings yet
- 12lect Nerve1 Summary2017Document4 pages12lect Nerve1 Summary2017Dan ChoiNo ratings yet
- 10lect Muscle1 Summary2017Document3 pages10lect Muscle1 Summary2017Dan ChoiNo ratings yet
- 08 Cartilage Summary2017Document2 pages08 Cartilage Summary2017Dan ChoiNo ratings yet
- 09lect Bone Summary2017Document3 pages09lect Bone Summary2017Dan ChoiNo ratings yet
- 07lect EpitheliumCTissue Summary2017Document4 pages07lect EpitheliumCTissue Summary2017Dan ChoiNo ratings yet
- 06lect ECM Summary2017Document5 pages06lect ECM Summary2017Dan ChoiNo ratings yet
- 03lect Cell2 Summary2017Document4 pages03lect Cell2 Summary2017Dan ChoiNo ratings yet
- 01lect Intro Summary2017Document2 pages01lect Intro Summary2017Dan ChoiNo ratings yet
- Entrep Bazaar Rating SheetDocument7 pagesEntrep Bazaar Rating SheetJupiter WhitesideNo ratings yet
- Chapin Columbus DayDocument15 pagesChapin Columbus Dayaspj13No ratings yet
- Togaf Open Group Business ScenarioDocument40 pagesTogaf Open Group Business Scenariohmh97No ratings yet
- Wwe SVR 2006 07 08 09 10 11 IdsDocument10 pagesWwe SVR 2006 07 08 09 10 11 IdsAXELL ENRIQUE CLAUDIO MENDIETANo ratings yet
- FPSCDocument15 pagesFPSCBABER SULTANNo ratings yet
- Narrative ReportDocument6 pagesNarrative ReportAlyssa Marie AsuncionNo ratings yet
- Outline - Criminal Law - RamirezDocument28 pagesOutline - Criminal Law - RamirezgiannaNo ratings yet
- Samsung LE26A457Document64 pagesSamsung LE26A457logik.huNo ratings yet
- Crime Scene Drawing January Incident 10501-10600Document100 pagesCrime Scene Drawing January Incident 10501-10600columbinefamilyrequest100% (2)
- What Is SCOPIC Clause - A Simple Overview - SailorinsightDocument8 pagesWhat Is SCOPIC Clause - A Simple Overview - SailorinsightJivan Jyoti RoutNo ratings yet
- Project TitleDocument15 pagesProject TitleadvikaNo ratings yet
- The BreakupDocument22 pagesThe BreakupAllison CreaghNo ratings yet
- 1634313583!Document24 pages1634313583!Joseph Sanchez TalusigNo ratings yet
- Reflection On Sumilao CaseDocument3 pagesReflection On Sumilao CaseGyrsyl Jaisa GuerreroNo ratings yet
- Account Statement From 1 Jan 2017 To 30 Jun 2017Document2 pagesAccount Statement From 1 Jan 2017 To 30 Jun 2017Ujjain mpNo ratings yet
- GemDocument135 pagesGemZelia GregoriouNo ratings yet
- Steel and Timber Design: Arch 415Document35 pagesSteel and Timber Design: Arch 415Glennson BalacanaoNo ratings yet
- Monastery in Buddhist ArchitectureDocument8 pagesMonastery in Buddhist ArchitectureabdulNo ratings yet
- SULTANS OF SWING - Dire Straits (Impresión)Document1 pageSULTANS OF SWING - Dire Straits (Impresión)fabio.mattos.tkd100% (1)
- ICONS+Character+Creator+2007+v0 73Document214 pagesICONS+Character+Creator+2007+v0 73C.M. LewisNo ratings yet
- Diane Mediano CareerinfographicDocument1 pageDiane Mediano Careerinfographicapi-344393975No ratings yet
- HTTP Parameter PollutionDocument45 pagesHTTP Parameter PollutionSpyDr ByTeNo ratings yet
- TestDocument56 pagesTestFajri Love PeaceNo ratings yet
- Weill Cornell Medicine International Tax QuestionaireDocument2 pagesWeill Cornell Medicine International Tax QuestionaireboxeritoNo ratings yet
- Nandurbar District S.E. (CGPA) Nov 2013Document336 pagesNandurbar District S.E. (CGPA) Nov 2013Digitaladda IndiaNo ratings yet
- MC Donald'sDocument30 pagesMC Donald'sAbdullahWaliNo ratings yet
- APA CitationsDocument9 pagesAPA CitationsIslamNo ratings yet
- 2024 01 31 StatementDocument4 pages2024 01 31 StatementAlex NeziNo ratings yet
- Industrial Visit Report Part 2Document41 pagesIndustrial Visit Report Part 2Navratan JagnadeNo ratings yet
- ITC Green Centre: Gurgaon, IndiaDocument19 pagesITC Green Centre: Gurgaon, IndiaAgastya Dasari100% (2)