You might also like
- An Overview of Research On Diseases of Cactus PearDocument6 pagesAn Overview of Research On Diseases of Cactus PearHIMANSHU BARWARNo ratings yet
- Sensitivity of Colletotrichum Species Responsible For Banana Anthracnose Disease To Some Fungicides Used in Postharvest Treatments in Côte D'ivoireDocument6 pagesSensitivity of Colletotrichum Species Responsible For Banana Anthracnose Disease To Some Fungicides Used in Postharvest Treatments in Côte D'ivoireIJEAB JournalNo ratings yet
- Merging Biotechnology With Biological Control: Banana Musa Tissue Culture Plants Enhanced by Endophytic FungiDocument7 pagesMerging Biotechnology With Biological Control: Banana Musa Tissue Culture Plants Enhanced by Endophytic FungiAkash DoiphodeNo ratings yet
- Ambeleetal 2012Document11 pagesAmbeleetal 2012Dércio MassangaieNo ratings yet
- Lasianthera Africana P. BEAUV Against Anopheles Gambiae (Diptera: Culicidae)Document7 pagesLasianthera Africana P. BEAUV Against Anopheles Gambiae (Diptera: Culicidae)International Journal of Innovative Science and Research TechnologyNo ratings yet
- Otto2018 Article BacterialCankerOfCherryTreesPrDocument12 pagesOtto2018 Article BacterialCankerOfCherryTreesPrFrancisca Beltrán GuzmánNo ratings yet
- Evaluation of The Prophylactic Activity of Ethanolic Extract of Ricinus Communis L. Against Plasmodium Berghei in MiceDocument11 pagesEvaluation of The Prophylactic Activity of Ethanolic Extract of Ricinus Communis L. Against Plasmodium Berghei in MiceUMYU Journal of Microbiology Research (UJMR)No ratings yet
- PE - Canker and Twig Dieback of Blueberry Caused by Pestalotiopsis Spp. and A Truncatella Sp. in ChileDocument8 pagesPE - Canker and Twig Dieback of Blueberry Caused by Pestalotiopsis Spp. and A Truncatella Sp. in ChileGabriela Scheinpflug BritoNo ratings yet
- 17 Ijasrapr2017017Document8 pages17 Ijasrapr2017017TJPRC PublicationsNo ratings yet
- Fusarium First Report 23Document3 pagesFusarium First Report 23api-676767477No ratings yet
- Incidence and Distribution of Citrus Tristeza Virus in Citrus Cultivars in Ibadan, Southwest NigeriaDocument4 pagesIncidence and Distribution of Citrus Tristeza Virus in Citrus Cultivars in Ibadan, Southwest NigeriaShailendra RajanNo ratings yet
- Vayssièresetal 2010 diversityfruitfliesCitrusBeninDocument17 pagesVayssièresetal 2010 diversityfruitfliesCitrusBeninperivou agoulaNo ratings yet
- Efficacy of Pulsed UV-Light For The Decontamination of Escherichia Coli O157:H7 and Salmonella Spp. On Raspberries and StrawberriesDocument7 pagesEfficacy of Pulsed UV-Light For The Decontamination of Escherichia Coli O157:H7 and Salmonella Spp. On Raspberries and StrawberriesFélix BaezNo ratings yet
- Protocol Optimisation For Micropropagation of Ethiopian YAMDocument15 pagesProtocol Optimisation For Micropropagation of Ethiopian YAMbahiran alwaysNo ratings yet
- Insecticidal Activity of A Destruxin-Containing Extract of Metarhizium Brunneum Against Ceratitis Capitata (Diptera: Tephritidae)Document11 pagesInsecticidal Activity of A Destruxin-Containing Extract of Metarhizium Brunneum Against Ceratitis Capitata (Diptera: Tephritidae)ahmredNo ratings yet
- Fita Et Al.2021.MolecularIdentification of White Mango Scale and Associated Natural Enemies In.... EthiopiaDocument14 pagesFita Et Al.2021.MolecularIdentification of White Mango Scale and Associated Natural Enemies In.... EthiopiaTemesgen FitaNo ratings yet
- Ekesietal 2009 Competitive DisplacementCcosyraBBinvades 81950Document11 pagesEkesietal 2009 Competitive DisplacementCcosyraBBinvades 81950perivou agoulaNo ratings yet
- Effect PhysiopathologiqueDocument9 pagesEffect PhysiopathologiqueJam Tog-Na Kami NicolasNo ratings yet
- Paper IJTDocument9 pagesPaper IJTdammini PremachandraNo ratings yet
- Larvicidal PropertyDocument6 pagesLarvicidal Propertychetykim padillo kimmayongNo ratings yet
- Study of Antagonistic Beneficial Microorganisms To Phytophtora Colocasiae Causal Agent of Taro Mildew Colocasia Esculenta L SchottDocument10 pagesStudy of Antagonistic Beneficial Microorganisms To Phytophtora Colocasiae Causal Agent of Taro Mildew Colocasia Esculenta L SchottLavile SakouvoguiNo ratings yet
- Isolation and Identification of Phytopathogenic Bacteria in Vegetable Crops in West Africa (Côte D'ivoire)Document11 pagesIsolation and Identification of Phytopathogenic Bacteria in Vegetable Crops in West Africa (Côte D'ivoire)dawit gNo ratings yet
- Shaltiel2015 - Control Tomato Leafminer Lepidoptera OpenField Tomatoes by Indigenous Natural Enemies OccurringDocument12 pagesShaltiel2015 - Control Tomato Leafminer Lepidoptera OpenField Tomatoes by Indigenous Natural Enemies Occurringeffi alfiani sidikNo ratings yet
- Breeding of Cocoa Trees (Theobroma Cacao L.) Resistantto Phytophthora Megakarya, Agent of Black Pod Disease in Cote DivoireDocument11 pagesBreeding of Cocoa Trees (Theobroma Cacao L.) Resistantto Phytophthora Megakarya, Agent of Black Pod Disease in Cote DivoireIJAR JOURNALNo ratings yet
- Parasites of Edible VegetablesDocument12 pagesParasites of Edible VegetablesVictor VinesNo ratings yet
- Inventory of Fungal Pathogens of Early Maturing Mango Varieties in The Kounkane Area, Southeast of SenegalDocument11 pagesInventory of Fungal Pathogens of Early Maturing Mango Varieties in The Kounkane Area, Southeast of SenegalOpenaccess Research paperNo ratings yet
- CoconutDocument37 pagesCoconutsautulhakimNo ratings yet
- Eng Journal 2Document9 pagesEng Journal 2Isnaeni RachmawatiNo ratings yet
- Plant Pathology - 2021 - Kimunye - Sources of Resistance To Pseudocercospora Fijiensis The Cause of Black Sigatoka inDocument14 pagesPlant Pathology - 2021 - Kimunye - Sources of Resistance To Pseudocercospora Fijiensis The Cause of Black Sigatoka inSandra Carolina Daza GachaNo ratings yet
- Effect of Temperature and Water Activity On in Vitro Germination of Monilinia SPPDocument8 pagesEffect of Temperature and Water Activity On in Vitro Germination of Monilinia SPPOana PotocNo ratings yet
- International Food Science and TechnologyDocument6 pagesInternational Food Science and Technologydeli liliaNo ratings yet
- Dry Heat Treatment of Andean Lupin Seed To Reduce Anthracnose InfectionDocument6 pagesDry Heat Treatment of Andean Lupin Seed To Reduce Anthracnose InfectionpepeNo ratings yet
- Nkya Vol212Document10 pagesNkya Vol212nephylymNo ratings yet
- Viljoen2019 Article BiologicalControlOfTheRoot-knoDocument8 pagesViljoen2019 Article BiologicalControlOfTheRoot-knonl14970No ratings yet
- Biological Control: Gang Fu, Siliang Huang, Yunfeng Ye, Yongguan Wu, Zhenlu Cen, Shanhai LinDocument10 pagesBiological Control: Gang Fu, Siliang Huang, Yunfeng Ye, Yongguan Wu, Zhenlu Cen, Shanhai LinRestrepo JuanNo ratings yet
- 10 1 1 866 5972 PDFDocument15 pages10 1 1 866 5972 PDFLinh NguyenNo ratings yet
- Proceso de Infección de Sigatoka in VivoDocument11 pagesProceso de Infección de Sigatoka in VivoCristhian JohnNo ratings yet
- Emma - Pdis 06 22 1350 SCDocument3 pagesEmma - Pdis 06 22 1350 SCOlajide Emmanuel OlorunfemiNo ratings yet
- Journal of Asia-Paci Fic Entomology: C. Somta, A. Winotai, P.A.C. OoiDocument4 pagesJournal of Asia-Paci Fic Entomology: C. Somta, A. Winotai, P.A.C. OoiFransiska VitaNo ratings yet
- Larvicidal Property of Sugar Apple (Annona Squamosa) and San Francisco (Codiaeum Variegatum) Against Yellow Fever Mosquito (Aedes Aegypti)Document4 pagesLarvicidal Property of Sugar Apple (Annona Squamosa) and San Francisco (Codiaeum Variegatum) Against Yellow Fever Mosquito (Aedes Aegypti)Euan VegaNo ratings yet
- Ivan Rwomushana, Sunday Ekesi, Callistus K.P.O. Ogol & Ian GordonDocument11 pagesIvan Rwomushana, Sunday Ekesi, Callistus K.P.O. Ogol & Ian Gordonperivou agoulaNo ratings yet
- Buku PerpajakanDocument10 pagesBuku PerpajakanSaadah 0210No ratings yet
- AjayiandOyedele-EVALUATIONOFAlliumsativumA LinnCRUDEEXTRACTSANDTrichodermaasperellumDocument8 pagesAjayiandOyedele-EVALUATIONOFAlliumsativumA LinnCRUDEEXTRACTSANDTrichodermaasperellumAjoke AdegayeNo ratings yet
- Pyrethroid Resistance in An Population From Uganda: Anopheles FunestusDocument8 pagesPyrethroid Resistance in An Population From Uganda: Anopheles Funestusibrahima1968No ratings yet
- Bacterial Diseases of Bananas and Enset Current State of Knowledge and Integrated Approaches Toward Sustainable ManagementDocument25 pagesBacterial Diseases of Bananas and Enset Current State of Knowledge and Integrated Approaches Toward Sustainable ManagementTheresa Anne MatunanNo ratings yet
- Bib OverviewDocument100 pagesBib OverviewMithun RajNo ratings yet
- As V17i2 4 PDFDocument10 pagesAs V17i2 4 PDFEbi KogaranaNo ratings yet
- Jee102 0144Document8 pagesJee102 0144Vicente MottaNo ratings yet
- ManuscriptDocument7 pagesManuscriptLuis NagueraNo ratings yet
- Jurnal Spoilage Guava Fruit - Mikpang PDFDocument6 pagesJurnal Spoilage Guava Fruit - Mikpang PDFdellavencaNo ratings yet
- Detection and Identification of The Coconut Lethal Yellowing Phytoplasma in Weeds Growing in Coconut Farms in C Te D IvoireDocument11 pagesDetection and Identification of The Coconut Lethal Yellowing Phytoplasma in Weeds Growing in Coconut Farms in C Te D IvoireQuevedodi IqdNo ratings yet
- Brontispa 2Document17 pagesBrontispa 2Earl Cadalin0% (1)
- Cactus Tissue CultureDocument8 pagesCactus Tissue CultureHüseyin Avni TokgözNo ratings yet
- Low-Dose Irradiation With Modified Atmosphere Packaging For Mango Against The Oriental Fruit Fly (Diptera: Tephritidae)Document6 pagesLow-Dose Irradiation With Modified Atmosphere Packaging For Mango Against The Oriental Fruit Fly (Diptera: Tephritidae)Haidar Ali AsshiddiqinNo ratings yet
- 68637-Article Text-142764-1-10-20110805Document9 pages68637-Article Text-142764-1-10-20110805Uroawuchi Eileen IhuomaNo ratings yet
- Biological Control: Abraha O. Abraham, Mark D. Laing, John P. BowerDocument7 pagesBiological Control: Abraha O. Abraham, Mark D. Laing, John P. BowernocrisdijoNo ratings yet
- Pdis 09 15 0974 ReDocument6 pagesPdis 09 15 0974 ReAbiel GutiérrezNo ratings yet
- Development of An in Vitro Culture System AdaptedDocument8 pagesDevelopment of An in Vitro Culture System AdaptedAndreina ZamoraNo ratings yet
- Agent Biology-The Potency of Endofit Fungi in Cocoa AsDocument6 pagesAgent Biology-The Potency of Endofit Fungi in Cocoa Asglenzi fizulmiNo ratings yet
- EPPO 2010 BactrocerazonatacontrolDocument6 pagesEPPO 2010 Bactrocerazonatacontrolperivou agoulaNo ratings yet
- Ekesietal 2009 Competitive DisplacementCcosyraBBinvades 81950Document11 pagesEkesietal 2009 Competitive DisplacementCcosyraBBinvades 81950perivou agoulaNo ratings yet
- Ekesi Et Al. 2016 Fruit Fly Research and Development in AfricaDocument776 pagesEkesi Et Al. 2016 Fruit Fly Research and Development in Africaperivou agoulaNo ratings yet
- Ekesietal 2007 Rearingbinvades 81950Document6 pagesEkesietal 2007 Rearingbinvades 81950perivou agoulaNo ratings yet
- Duycketal 2006 nichepartitioningTephritidaeReunionDocument9 pagesDuycketal 2006 nichepartitioningTephritidaeReunionperivou agoulaNo ratings yet
- Farmers' Preferences For The Design of Fruit Fly Pest-Free Area 2015Document12 pagesFarmers' Preferences For The Design of Fruit Fly Pest-Free Area 2015perivou agoulaNo ratings yet
- Ekesietal - 2006 - LifehistorybinvadensDocument8 pagesEkesietal - 2006 - Lifehistorybinvadensperivou agoulaNo ratings yet
- DeMeyeretal - 2002 - Annotated Check List of Host PlantsDocument87 pagesDeMeyeretal - 2002 - Annotated Check List of Host Plantsperivou agoulaNo ratings yet
- Duycketal 2007 Kselection&invaders 82033Document9 pagesDuycketal 2007 Kselection&invaders 82033perivou agoulaNo ratings yet
- Ivan Rwomushana, Sunday Ekesi, Callistus K.P.O. Ogol & Ian GordonDocument11 pagesIvan Rwomushana, Sunday Ekesi, Callistus K.P.O. Ogol & Ian Gordonperivou agoulaNo ratings yet
- De Villiersetal 2012 CLIMEXCeratitisrosaDocument14 pagesDe Villiersetal 2012 CLIMEXCeratitisrosaperivou agoulaNo ratings yet
- Sagrietal - 2014 - Spinosad Resistance in OliveflyDocument20 pagesSagrietal - 2014 - Spinosad Resistance in Oliveflyperivou agoulaNo ratings yet
- De Meyeretal 2013 WestafricanfruitfliesDocument17 pagesDe Meyeretal 2013 Westafricanfruitfliesperivou agoulaNo ratings yet
- Vargas ManagmentDocument22 pagesVargas Managmentperivou agoulaNo ratings yet
- Vayssièresetal 2010 BiocontrolParasitoidsDocument9 pagesVayssièresetal 2010 BiocontrolParasitoidsperivou agoulaNo ratings yet
- Schuler Et Al 2016 Molecular EcologyDocument40 pagesSchuler Et Al 2016 Molecular Ecologyperivou agoulaNo ratings yet
- Vayssieresetal 2007 cucurbitaeWAfricaDocument6 pagesVayssieresetal 2007 cucurbitaeWAfricaperivou agoulaNo ratings yet
- A New Bactrocera Species in Benin Among Mango Fruit Fly (Diptera: Tephritidae) SpeciesDocument7 pagesA New Bactrocera Species in Benin Among Mango Fruit Fly (Diptera: Tephritidae) Speciesperivou agoulaNo ratings yet
- Geurtsetal 2012 Altitudinaltransect-RedDocument18 pagesGeurtsetal 2012 Altitudinaltransect-Redperivou agoulaNo ratings yet
- Takanoetal - 2014 - Bactrocera Latifrons Survival TemperatureDocument9 pagesTakanoetal - 2014 - Bactrocera Latifrons Survival Temperatureperivou agoulaNo ratings yet
- Vayssièresetal 2010 diversityfruitfliesCitrusBeninDocument17 pagesVayssièresetal 2010 diversityfruitfliesCitrusBeninperivou agoulaNo ratings yet
- Vayssieresetal 2009 FruitflycorrelationDocument12 pagesVayssieresetal 2009 Fruitflycorrelationperivou agoulaNo ratings yet
- Vayssièresetal 2010 BiocontrolParasitoidsDocument9 pagesVayssièresetal 2010 BiocontrolParasitoidsperivou agoulaNo ratings yet
- Effectiveness of Spinosad Bait Sprays (GF-120) in Controlling Mango-Infesting Fruit Flies (Diptera: Tephritidae) in BeninDocument7 pagesEffectiveness of Spinosad Bait Sprays (GF-120) in Controlling Mango-Infesting Fruit Flies (Diptera: Tephritidae) in Beninperivou agoulaNo ratings yet
- De Meyeretal - 2012 - TephritidaecomoresDocument10 pagesDe Meyeretal - 2012 - Tephritidaecomoresperivou agoulaNo ratings yet
- Giovanni Benelli Fighting of Fruits Flies 2014Document16 pagesGiovanni Benelli Fighting of Fruits Flies 2014perivou agoulaNo ratings yet
- Farmers' Preferences For The Design of Fruit Fly Pest-Free Area 2015Document12 pagesFarmers' Preferences For The Design of Fruit Fly Pest-Free Area 2015perivou agoulaNo ratings yet
- EPPO 2010 BactrocerazonatacontrolDocument6 pagesEPPO 2010 Bactrocerazonatacontrolperivou agoulaNo ratings yet
- Duycketal 2006 nichepartitioningTephritidaeReunionDocument9 pagesDuycketal 2006 nichepartitioningTephritidaeReunionperivou agoulaNo ratings yet
- Asphyxia: DR Kaleem KhanDocument81 pagesAsphyxia: DR Kaleem KhanShafira WidiaNo ratings yet
- A Multi-Dimensional Model of Dysfunctional BreathiDocument4 pagesA Multi-Dimensional Model of Dysfunctional Breathimari_kundera19No ratings yet
- Heat Stress in PlantsDocument22 pagesHeat Stress in PlantsPao GabrielNo ratings yet
- In The Devils TeethDocument129 pagesIn The Devils TeethAprilia Kartika PutriNo ratings yet
- Bleuler Psychology PDFDocument666 pagesBleuler Psychology PDFRicardo Jacobsen Gloeckner100% (2)
- Carlson (7E) : Powerpoint Lecture OutlineDocument26 pagesCarlson (7E) : Powerpoint Lecture Outlinechat gazaNo ratings yet
- Basic Anatomy and Physiology 2Document6 pagesBasic Anatomy and Physiology 2Mohd HadhriNo ratings yet
- Natures Hidden Health MiracleDocument38 pagesNatures Hidden Health MiracleGorneau100% (2)
- G11 Biology EnglishDocument7 pagesG11 Biology Englishahmed aluNo ratings yet
- Airway Problems in PACUDocument35 pagesAirway Problems in PACUIndrani HemantkumarNo ratings yet
- Molecular Biology ExperimentDocument6 pagesMolecular Biology ExperimentWalwin Hare100% (1)
- Urologic Surgery BOOK Sabiston Textbook of Surgery, Twentieth Edition Townsend, Courtney M., JR., Beauchamp, R. Daniel Evers, B. Mark, Mattox, Kenneth LDocument39 pagesUrologic Surgery BOOK Sabiston Textbook of Surgery, Twentieth Edition Townsend, Courtney M., JR., Beauchamp, R. Daniel Evers, B. Mark, Mattox, Kenneth Loeij stevanus WijayaNo ratings yet
- Cerebellum & Brain-Stem AnatomyDocument23 pagesCerebellum & Brain-Stem AnatomyMuhammad ShahzaibNo ratings yet
- Ecg en LabviewDocument61 pagesEcg en LabviewHarold David Gil MuñozNo ratings yet
- PACHECO - EXERCISE NO.1-The CellDocument3 pagesPACHECO - EXERCISE NO.1-The CellMaria Brooklyn PachecoNo ratings yet
- 4 - Ditta - BIO173 - Unity of Life - 20204Document13 pages4 - Ditta - BIO173 - Unity of Life - 20204John ChoiNo ratings yet
- Tactical BreathingDocument3 pagesTactical Breathingsergio661204No ratings yet
- ISMST Shockwave Medical Papers 2012Document24 pagesISMST Shockwave Medical Papers 2012Dr-Gehad Samy Halwagy100% (1)
- Chakras and The Lord's ParayerDocument10 pagesChakras and The Lord's Parayermoura_beth32260% (1)
- Anatomia de BufalosDocument24 pagesAnatomia de BufalosTrilce ClaroNo ratings yet
- Enzyme Catalysis. FOXIT - Not Different, Just Better NAT91 Sin ReferenciasDocument6 pagesEnzyme Catalysis. FOXIT - Not Different, Just Better NAT91 Sin Referenciaskiper.valNo ratings yet
- Bio 130 Chapter 4 NotesDocument12 pagesBio 130 Chapter 4 Notesgilissa100% (1)
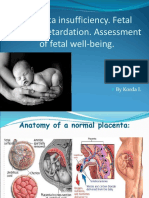
- Placenta InsufficiencyDocument39 pagesPlacenta InsufficiencyslyfoxkittyNo ratings yet
- Introduction To Neuroscience: Systems Neuroscience - Concepts and MethodsDocument64 pagesIntroduction To Neuroscience: Systems Neuroscience - Concepts and MethodsAldo Cesar BCNo ratings yet
- Polymers in Medicine and Surgery 1975 PDFDocument336 pagesPolymers in Medicine and Surgery 1975 PDFVidvendu GuptaNo ratings yet
- Aits Neet Grand Test - 23 Paper Key (04-05-2023)Document10 pagesAits Neet Grand Test - 23 Paper Key (04-05-2023)vulurakashsharma2005No ratings yet
- Genetics in Blood BankingDocument47 pagesGenetics in Blood BankingMandy A. Delfin100% (3)
- PHARMACOLOGYDocument36 pagesPHARMACOLOGYjanr123456100% (1)
- It All Began With A Love BiteDocument17 pagesIt All Began With A Love Biteapi-262208234No ratings yet
- Calf Rearing Practice - ManualDocument33 pagesCalf Rearing Practice - ManualJose DeJesus Bustamante GuerreroNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)