You might also like
- Delta Opioid Receptor Pharmacology and Therapeutic ApplicationsFrom EverandDelta Opioid Receptor Pharmacology and Therapeutic ApplicationsEmily M. JutkiewiczNo ratings yet
- Articulo IVICDocument11 pagesArticulo IVICMuôngNo ratings yet
- Procaine and Procaine-Base-Infusion - A Review of The Safety and Fields of Application After Twenty Years of UseDocument7 pagesProcaine and Procaine-Base-Infusion - A Review of The Safety and Fields of Application After Twenty Years of UseJuan Sebastián Cerón OtáloraNo ratings yet
- 2501 Depression and Mental HeDocument84 pages2501 Depression and Mental Herichardong lauNo ratings yet
- Essential Phospholipids in Fatty Liver - Scientific Update CEG-9-105 2016Document13 pagesEssential Phospholipids in Fatty Liver - Scientific Update CEG-9-105 2016Adriel MirtoNo ratings yet
- Expression of Opioid Receptors in Cells of The Immune SystemDocument13 pagesExpression of Opioid Receptors in Cells of The Immune Systemulfi septawartiningtiasNo ratings yet
- Anti EpilepticDocument13 pagesAnti EpilepticArbiati RahmanNo ratings yet
- AnandDocument12 pagesAnandre septian IlhamsyahNo ratings yet
- Treatment-Resistant Obsessive-Compulsive Disorder (OCD) - Current UnderstandingDocument21 pagesTreatment-Resistant Obsessive-Compulsive Disorder (OCD) - Current UnderstandingRapapportNo ratings yet
- 2014 Article 605Document10 pages2014 Article 605Fi NoNo ratings yet
- Hyperbaric Oxygenation Therapy: Molecular Mechanisms and Clinical ApplicationsFrom EverandHyperbaric Oxygenation Therapy: Molecular Mechanisms and Clinical ApplicationsNariyoshi ShinomiyaNo ratings yet
- Zhang, Yu, Zhang - ARA 290 Relieves Pathophysiological Pain by Targeting TRPV1 Channel - Integration Between Immune System and NociceptionDocument27 pagesZhang, Yu, Zhang - ARA 290 Relieves Pathophysiological Pain by Targeting TRPV1 Channel - Integration Between Immune System and Nociception朗博CuauhtlatoaNo ratings yet
- AIJPMS - Volume 3 - Issue 1 - Pages 15-31Document17 pagesAIJPMS - Volume 3 - Issue 1 - Pages 15-31Eman MahmoudNo ratings yet
- IJPSRDocument8 pagesIJPSRNithin VijayakumarNo ratings yet
- Pharmacokinetic-Pharmacodynamic Analysis On Inflammation Rat Model After Oral Administration of Huang Lian Jie Du DecoctionDocument17 pagesPharmacokinetic-Pharmacodynamic Analysis On Inflammation Rat Model After Oral Administration of Huang Lian Jie Du Decoctionivo maideNo ratings yet
- IJN 76092 Nanoformulation of D A Tocopheryl Polyethylene Glycol 1000 S 021715Document12 pagesIJN 76092 Nanoformulation of D A Tocopheryl Polyethylene Glycol 1000 S 021715MuthmainnahNo ratings yet
- The in Vitro Receptor Profile of Rotigotine: A New Agent For The Treatment of ParkinsonDocument14 pagesThe in Vitro Receptor Profile of Rotigotine: A New Agent For The Treatment of ParkinsonPietro BracagliaNo ratings yet
- Neuroscience Letters: Laura Monni, Filippo Ghezzi, Silvia Corsini, Andrea NistriDocument6 pagesNeuroscience Letters: Laura Monni, Filippo Ghezzi, Silvia Corsini, Andrea NistriardhiNo ratings yet
- Pharmacology Concept BookDocument357 pagesPharmacology Concept Bookram dheerNo ratings yet
- 11 Iajps11072017Document7 pages11 Iajps11072017Baru Chandrasekhar RaoNo ratings yet
- LASTDocument10 pagesLASTAgus RiyantoNo ratings yet
- Research ArticleDocument9 pagesResearch ArticleRajesh KumarNo ratings yet
- Animals 11 02199Document10 pagesAnimals 11 02199Aridai RJNo ratings yet
- JKSHP033 01 05Document12 pagesJKSHP033 01 05dennystefanus03No ratings yet
- Research Article Pleurotus Eryngii Ameliorates Lipopolysaccharide-InducedDocument7 pagesResearch Article Pleurotus Eryngii Ameliorates Lipopolysaccharide-InducedkierkegardNo ratings yet
- Novel Therapy For Hearing Loss: Delivery of Insulin-Like Growth Factor 1 To The Cochlea Using Gelatin HydrogelDocument6 pagesNovel Therapy For Hearing Loss: Delivery of Insulin-Like Growth Factor 1 To The Cochlea Using Gelatin HydrogelHinami TenukiNo ratings yet
- Interaction of Quercetin of Onion With Axon Guidance Protein Receptor, NRP-1 Plays Important Role in Cancer Treatment: An in Silico ApproachDocument8 pagesInteraction of Quercetin of Onion With Axon Guidance Protein Receptor, NRP-1 Plays Important Role in Cancer Treatment: An in Silico Approachkhangkhang151203No ratings yet
- European Journal of Medicinal Chemistry: Original ArticleDocument11 pagesEuropean Journal of Medicinal Chemistry: Original ArticleRômuloNo ratings yet
- Upregulation of Opioid ReceptorsDocument26 pagesUpregulation of Opioid ReceptorsbrightfutureNo ratings yet
- The Clinical Pharmacology and Pharmacokinetics of Ulipristal Acetate For The Treatment of Uterine FibroidsDocument8 pagesThe Clinical Pharmacology and Pharmacokinetics of Ulipristal Acetate For The Treatment of Uterine FibroidsZakia DrajatNo ratings yet
- The Pharmacokinetics of Long-Acting Antipsychotic Medications. 9 (2011)Document8 pagesThe Pharmacokinetics of Long-Acting Antipsychotic Medications. 9 (2011)SolomonNo ratings yet
- Research Article: Acupuncture Attenuated Inflammation and Inhibited Th17 and Treg Activity in Experimental AsthmaDocument8 pagesResearch Article: Acupuncture Attenuated Inflammation and Inhibited Th17 and Treg Activity in Experimental AsthmaAdán LópezNo ratings yet
- Mechanisms of Hypoxia Signalling: New Implications For NephrologyDocument19 pagesMechanisms of Hypoxia Signalling: New Implications For Nephrologyabdul88fNo ratings yet
- pdf-3Document6 pagespdf-3Mohammad SafiqulNo ratings yet
- Ni Hms 904626Document15 pagesNi Hms 904626omar gonzalezNo ratings yet
- Ligustrazine Phosphate Ethosomes For Treatment of Alzheimer's Disease, in Vitro and in Animal Model StudiesDocument8 pagesLigustrazine Phosphate Ethosomes For Treatment of Alzheimer's Disease, in Vitro and in Animal Model Studiesebooks list1No ratings yet
- Pharmacology of AutacoidsDocument13 pagesPharmacology of AutacoidsInocenteNo ratings yet
- AripripazoleDocument13 pagesAripripazoleOneng IfayaniNo ratings yet
- Rocuronio Nao Influencia No PSIDocument7 pagesRocuronio Nao Influencia No PSIbrasilvilermandoNo ratings yet
- Research ArticleDocument8 pagesResearch ArticleelinNo ratings yet
- Bioequivalence Study of Two Loperamide HydrochloriDocument5 pagesBioequivalence Study of Two Loperamide HydrochloriLoRna Nu'aNo ratings yet
- Formulation and Evaluation of Time Dependendt Release of Montelukast Tablets by Using Mini Tablet TechnologyDocument13 pagesFormulation and Evaluation of Time Dependendt Release of Montelukast Tablets by Using Mini Tablet TechnologyBaru Chandrasekhar RaoNo ratings yet
- Short Term Induction of Lps Damages The Hippocampus Gyrus Through Apoptosis On Glial Cells MiceDocument9 pagesShort Term Induction of Lps Damages The Hippocampus Gyrus Through Apoptosis On Glial Cells MiceGlobal Research and Development ServicesNo ratings yet
- Primary Literature Review PosterDocument1 pagePrimary Literature Review PosterZach PipkinNo ratings yet
- 1 s2.0 S1347861317300269 MainDocument7 pages1 s2.0 S1347861317300269 MainMadhanNo ratings yet
- Ozone Therapy Ameliorates Tubulointerstitial Inflammation by Regulating TLR4 in Adenine Induced CKD RatsDocument10 pagesOzone Therapy Ameliorates Tubulointerstitial Inflammation by Regulating TLR4 in Adenine Induced CKD RatsBoby FaesalNo ratings yet
- 6578 FullDocument12 pages6578 Fullシン イNo ratings yet
- Al4 PDFDocument6 pagesAl4 PDFDicNo ratings yet
- Physiology of WDDocument16 pagesPhysiology of WDphungoNo ratings yet
- Deshmukh Et Al 2021 Palmitoylethanolamide Prenatal Developmental Toxicity Study in RatsDocument10 pagesDeshmukh Et Al 2021 Palmitoylethanolamide Prenatal Developmental Toxicity Study in RatssubhamNo ratings yet
- Drug Anim, AlDocument5 pagesDrug Anim, AlakondisNo ratings yet
- DonepezilAreview PDFDocument19 pagesDonepezilAreview PDFDesi PermatasariNo ratings yet
- Optimization of Chronomodulated Delivery System Coated With A Blend of Ethyl Cellulose and Eudragit L100 by Central Composite Design: in Vitro and in Vivo EvaluationDocument11 pagesOptimization of Chronomodulated Delivery System Coated With A Blend of Ethyl Cellulose and Eudragit L100 by Central Composite Design: in Vitro and in Vivo EvaluationIIrsandi JohanNo ratings yet
- Journal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From MitochondrialDocument13 pagesJournal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From Mitochondrialjalewa.jaishreeNo ratings yet
- Helium Preconditioning Protects Against Neonatal Hypoxia-IschemiaDocument7 pagesHelium Preconditioning Protects Against Neonatal Hypoxia-IschemiaJohn BiniarisNo ratings yet
- The Gut-Brain Interaction in Opioid Tolerance: SciencedirectDocument5 pagesThe Gut-Brain Interaction in Opioid Tolerance: SciencedirectNatalia SantacruzNo ratings yet
- 1a. 24609742 Neurobiology of PainDocument6 pages1a. 24609742 Neurobiology of PainJason HillNo ratings yet
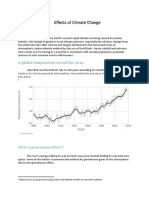
- Effects of Climate ChangeDocument3 pagesEffects of Climate Changejiofjij100% (1)
- LET General Math ReviewerDocument7 pagesLET General Math ReviewerMarco Rhonel Eusebio100% (1)
- World's Standard Model G6A!: Low Signal RelayDocument9 pagesWorld's Standard Model G6A!: Low Signal RelayEgiNo ratings yet
- Twilight PrincessDocument49 pagesTwilight PrincessHikari DiegoNo ratings yet
- TM-8000 HD Manual PDFDocument37 pagesTM-8000 HD Manual PDFRoxana BirtumNo ratings yet
- Recipes From The Perfect Scoop by David LebovitzDocument10 pagesRecipes From The Perfect Scoop by David LebovitzThe Recipe Club100% (7)
- Basic Geriatric Nursing 6th Edition Williams Test BankDocument10 pagesBasic Geriatric Nursing 6th Edition Williams Test Bankmaryrodriguezxsntrogkwd100% (49)
- Anxiety Disorders - Causes, Types, Symptoms, & TreatmentsDocument5 pagesAnxiety Disorders - Causes, Types, Symptoms, & Treatmentsrehaan662No ratings yet
- Catalogo Aesculap PDFDocument16 pagesCatalogo Aesculap PDFHansNo ratings yet
- Cathodic Protection Catalog - New 8Document1 pageCathodic Protection Catalog - New 8dhineshNo ratings yet
- Updated Factory Profile of Aleya Apparels LTDDocument25 pagesUpdated Factory Profile of Aleya Apparels LTDJahangir Hosen0% (1)
- Tugas 2-TRK Lanjut Kelompok 3 Andre-Arief-IstiaDocument18 pagesTugas 2-TRK Lanjut Kelompok 3 Andre-Arief-IstiaAndre Fahriz Perdana HarahapNo ratings yet
- Schneider Pressure Switch XMLDocument2 pagesSchneider Pressure Switch XMLhaoNo ratings yet
- Design and Development of Automatic Pneumatic Jack in Four Wheeler Ijariie5374Document5 pagesDesign and Development of Automatic Pneumatic Jack in Four Wheeler Ijariie5374Saravanan ViswakarmaNo ratings yet
- Hydraulic Fan Speed - Test and AdjustDocument12 pagesHydraulic Fan Speed - Test and Adjustsyed ahmedNo ratings yet
- Column, Slab, Footing and Wall Footing Foundations: Class A MixingDocument47 pagesColumn, Slab, Footing and Wall Footing Foundations: Class A MixingGioharry Nul PanambulanNo ratings yet
- Guia de CondensadoresDocument193 pagesGuia de CondensadoresPaola Segura CorreaNo ratings yet
- Bomba MixerDocument2 pagesBomba MixerBinelli79No ratings yet
- SDHI18 - Komparativna Analiza Primene Vodostana I Sinhronih Regulatora TurbinaDocument13 pagesSDHI18 - Komparativna Analiza Primene Vodostana I Sinhronih Regulatora TurbinaAleksandar PetkovicNo ratings yet
- Harmonics PatternsDocument4 pagesHarmonics PatternsIzzadAfif1990No ratings yet
- Passenger Lift Alert - Health and Safety AuthorityDocument4 pagesPassenger Lift Alert - Health and Safety AuthorityReginald MaswanganyiNo ratings yet
- Arduino Oscilloscope ProjectDocument12 pagesArduino Oscilloscope ProjectSathya Narayan100% (1)
- Worksheet Series 5: Answer Any 3 Questions From 1 To 6. (2 Score Each)Document6 pagesWorksheet Series 5: Answer Any 3 Questions From 1 To 6. (2 Score Each)AswithNo ratings yet
- G.S 5Document2 pagesG.S 5Jamsher BalochNo ratings yet
- American BreakfastDocument4 pagesAmerican BreakfastHamilton Valenzuela ChipongianNo ratings yet
- Halfen Cast-In Channels: HTA-CE 50/30P HTA-CE 40/22PDocument92 pagesHalfen Cast-In Channels: HTA-CE 50/30P HTA-CE 40/22PTulusNo ratings yet
- Food Taste Panel Evaluation Form 2Document17 pagesFood Taste Panel Evaluation Form 2Akshat JainNo ratings yet
- CheckList For Checking of Drawings-R1Document4 pagesCheckList For Checking of Drawings-R1jatin kalraNo ratings yet
- FRP Handrail Fittings CatalogDocument6 pagesFRP Handrail Fittings CatalogAl Adel MorenoNo ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet