You might also like
- Microbial Glycobiology: Structures, Relevance and ApplicationsFrom EverandMicrobial Glycobiology: Structures, Relevance and ApplicationsNo ratings yet
- Regulation of Peptidoglycan SynthesisDocument15 pagesRegulation of Peptidoglycan SynthesisVictor HugoNo ratings yet
- Printed in Great Britain. All Rights Reserved, ©: Biotech. Adv. Vol. 9, Pp. 217-240,1991Document24 pagesPrinted in Great Britain. All Rights Reserved, ©: Biotech. Adv. Vol. 9, Pp. 217-240,1991sebastian floresNo ratings yet
- Polymers 15 03481 With CoverDocument20 pagesPolymers 15 03481 With Coverlidya osNo ratings yet
- International Journal of Medical MicrobiologyDocument10 pagesInternational Journal of Medical MicrobiologyArthur Zanetti Nunes FernandesNo ratings yet
- Endotoxin and Disease in Food AnimalsDocument8 pagesEndotoxin and Disease in Food Animalstaner_soysuren100% (1)
- Guenin-Mac - Et Al-2009-Transboundary and Emerging DiseasesDocument14 pagesGuenin-Mac - Et Al-2009-Transboundary and Emerging DiseasesBibiana Chavarro PortilloNo ratings yet
- Structure, Biochemistry and Mechanism of Action of Glycopeptide AntibioticsDocument8 pagesStructure, Biochemistry and Mechanism of Action of Glycopeptide Antibioticsl4vfeaokf5No ratings yet
- Biochimica Et Biophysica Acta: Richard M. Epand, Chelsea Walker, Raquel F. Epand, Nathan A. MagarveyDocument8 pagesBiochimica Et Biophysica Acta: Richard M. Epand, Chelsea Walker, Raquel F. Epand, Nathan A. MagarveyAgung RahmadaniNo ratings yet
- Bacterial StructureDocument72 pagesBacterial StructureKrittika ChatterjeeNo ratings yet
- Microbiology 2Document15 pagesMicrobiology 2husseinNo ratings yet
- Resistensi Dan Cara Kerja AntiboticsDocument6 pagesResistensi Dan Cara Kerja AntiboticsChristiadi karlin12No ratings yet
- Members of The Microbial WorldDocument99 pagesMembers of The Microbial WorldShermin EeNo ratings yet
- 2014 Microbial Cell Factories ReviewDocument24 pages2014 Microbial Cell Factories ReviewXgfyfgyhNo ratings yet
- Aml 170423Document35 pagesAml 170423Aleksa SesicNo ratings yet
- Action and Resistance Mechanisms of Antibiotics: A Guide For CliniciansDocument6 pagesAction and Resistance Mechanisms of Antibiotics: A Guide For CliniciansAlejandra SánchezNo ratings yet
- Action and Resistance Mechanisms of Antibiotics - A Guide For CliniciansDocument6 pagesAction and Resistance Mechanisms of Antibiotics - A Guide For CliniciansJonatas SilvaNo ratings yet
- MCB 120 Exam 1ADocument66 pagesMCB 120 Exam 1AMark TolentinoNo ratings yet
- 2019 - The Genetics and Biochemistry of Cell Wall Structure and Synthesis in N CrassaDocument18 pages2019 - The Genetics and Biochemistry of Cell Wall Structure and Synthesis in N CrassaskbucherNo ratings yet
- Bacteria and Bacterial Diseases Lecture NotesDocument8 pagesBacteria and Bacterial Diseases Lecture NotesPowell KitagwaNo ratings yet
- Biochimica Et Biophysica Acta: Joel Selkrig, Denisse L. Leyton, Chaille T. Webb, Trevor LithgowDocument9 pagesBiochimica Et Biophysica Acta: Joel Selkrig, Denisse L. Leyton, Chaille T. Webb, Trevor LithgowalkatsumiNo ratings yet
- BIO350 BacStrucDocument65 pagesBIO350 BacStrucPrincewill SeiyefaNo ratings yet
- Bacterial Cell Structure FunctionsDocument27 pagesBacterial Cell Structure FunctionsFRANCIS PAOLO CUADRANo ratings yet
- 1 s2.0 S2211926422001576 MainDocument8 pages1 s2.0 S2211926422001576 MainAnthea TranNo ratings yet
- Chemistry and Histochemistry of Gram Staining of Dyes On Bacterial PeptidoglycanDocument46 pagesChemistry and Histochemistry of Gram Staining of Dyes On Bacterial PeptidoglycanJordi FrNo ratings yet
- Antibiotics 09 00553 PDFDocument21 pagesAntibiotics 09 00553 PDFGepetto ArtsNo ratings yet
- Lipid Mediated Antimicrobial Resistance: A Phantom Menace or A New Hope?Document18 pagesLipid Mediated Antimicrobial Resistance: A Phantom Menace or A New Hope?d99452727No ratings yet
- HHS Public Access: The Mycobacterium Tuberculosis Capsule: A Cell Structure With Key Implications in PathogenesisDocument34 pagesHHS Public Access: The Mycobacterium Tuberculosis Capsule: A Cell Structure With Key Implications in PathogenesisGrace Meigawati IwantoNo ratings yet
- Reviews the dynamic cell envelope of Mycobacterium tuberculosisDocument13 pagesReviews the dynamic cell envelope of Mycobacterium tuberculosisJose Maria RojasNo ratings yet
- Tissue Degradación PeptidosDocument13 pagesTissue Degradación PeptidosAngel Escandón VillaNo ratings yet
- Estructura y Arquitectura Del PeptidoglucanoDocument19 pagesEstructura y Arquitectura Del PeptidoglucanoOleggValVilNo ratings yet
- Mod 2Document24 pagesMod 2The GreatNo ratings yet
- Review The Short Proline-Rich Antibacterial Peptide Family: Cellular and Molecular Life SciencesDocument13 pagesReview The Short Proline-Rich Antibacterial Peptide Family: Cellular and Molecular Life Sciencesmutyarsih oryza sativaNo ratings yet
- Bacterial Cell StructureDocument9 pagesBacterial Cell StructureManWol JangNo ratings yet
- Cells: Perspectives For Glyco-Engineering of Recombinant Biopharmaceuticals From MicroalgaeDocument26 pagesCells: Perspectives For Glyco-Engineering of Recombinant Biopharmaceuticals From MicroalgaeVaibhav LonganiNo ratings yet
- Chapter 3 MIC260 PDFDocument36 pagesChapter 3 MIC260 PDFWan AiniNo ratings yet
- Bacterial MorphologyDocument21 pagesBacterial MorphologyGaspar SantosNo ratings yet
- Re-Evaluating The Importance of Carbohydrates As Regenerative BiomaterialsDocument12 pagesRe-Evaluating The Importance of Carbohydrates As Regenerative BiomaterialsJessica Tatiana Feo ZamoraNo ratings yet
- ART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansDocument16 pagesART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansHECTORIBZAN ACERO SANDOVALNo ratings yet
- Bacterial StructureDocument31 pagesBacterial StructureSaid AbdelaNo ratings yet
- Protein Sorting To The Cell Wall Envelope of Gram-Positive BacteriaDocument10 pagesProtein Sorting To The Cell Wall Envelope of Gram-Positive BacteriaDipankar RoyNo ratings yet
- Nihms 1831105Document49 pagesNihms 1831105Estefany Malpartida PalominoNo ratings yet
- Promising natural antimicrobial strategies for food biopreservationDocument10 pagesPromising natural antimicrobial strategies for food biopreservationTran Thach Uyen NhiNo ratings yet
- BST 49 915Document17 pagesBST 49 915bilou hertNo ratings yet
- Burkhart 2003Document3 pagesBurkhart 2003Trustia RizqandaruNo ratings yet
- Micro para 1Document4 pagesMicro para 1Reselle EspirituNo ratings yet
- DM-1 - Lesson 11Document10 pagesDM-1 - Lesson 11Amit GodaraNo ratings yet
- Review Molecular Interactions in BiofilmsDocument13 pagesReview Molecular Interactions in Biofilmsnaima aminaNo ratings yet
- Jof 08 00103 v2Document19 pagesJof 08 00103 v2tomsiriNo ratings yet
- Bacterial Cell Wall: Dr. B.Vijayasree 1 Year Post-Graduate SMC, VjaDocument42 pagesBacterial Cell Wall: Dr. B.Vijayasree 1 Year Post-Graduate SMC, Vjavijayasree bavireddyNo ratings yet
- Immunomodulation and Anti-Cancer Activity of Polysaccharide-Protein ComplexesDocument15 pagesImmunomodulation and Anti-Cancer Activity of Polysaccharide-Protein ComplexesPriyankar MajiNo ratings yet
- Bacterial Structure 1 1 2Document41 pagesBacterial Structure 1 1 2Nour MohammedNo ratings yet
- Antimicrobial Enzymes: Application and Future Potential in The Food IndustryDocument7 pagesAntimicrobial Enzymes: Application and Future Potential in The Food IndustryMauhibahYumnaNo ratings yet
- An in Vitro Cytotoxicity of A Novel PH Sensitive Lecti - 2021 - Journal of DrugDocument14 pagesAn in Vitro Cytotoxicity of A Novel PH Sensitive Lecti - 2021 - Journal of DrugRiki WijayaNo ratings yet
- Cell Walls of Prokaryotes - Boundless MicrobiologyDocument15 pagesCell Walls of Prokaryotes - Boundless MicrobiologyVy VyNo ratings yet
- Epidermidis Biofilms: Activity of Gallidermin On Staphylococcus Aureus and StaphylococcusDocument7 pagesEpidermidis Biofilms: Activity of Gallidermin On Staphylococcus Aureus and StaphylococcusnimertaNo ratings yet
- Vipin4C GlutamicumDocument16 pagesVipin4C GlutamicumerdilaputridivaNo ratings yet
- Bacterial ClassificationDocument5 pagesBacterial ClassificationShai KateNo ratings yet
- Lignocellulose Degradation Mechanisms Across The Tree of LifeDocument12 pagesLignocellulose Degradation Mechanisms Across The Tree of LifeAlonsoNo ratings yet
- Membrane BiochemistryDocument436 pagesMembrane BiochemistryJürgen Nicholas Schwarze100% (13)
- Laporan Praktikum Biofarmasetika 1 Percobaan 1: Optimasi Metode Analisa ObatDocument9 pagesLaporan Praktikum Biofarmasetika 1 Percobaan 1: Optimasi Metode Analisa ObatRizky Bachrul AlamNo ratings yet
- Salt Salt Analysis - 6 ZN (CO3) 2Document2 pagesSalt Salt Analysis - 6 ZN (CO3) 2mystical moonbeamNo ratings yet
- Separation Processes-II Adsorption and Fixed-Bed SeparationsDocument9 pagesSeparation Processes-II Adsorption and Fixed-Bed SeparationsAyush MehtaNo ratings yet
- Request Bearing Pad Test UrgentlyDocument1 pageRequest Bearing Pad Test UrgentlyShoyeeb AhmedNo ratings yet
- Thermal Processing Food-Related Toxicants: A Review: Critical Reviews in Food Science and NutritionDocument19 pagesThermal Processing Food-Related Toxicants: A Review: Critical Reviews in Food Science and NutritionRIFQI FAYYADH ANSARNo ratings yet
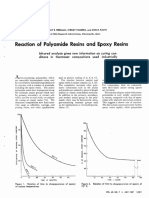
- Peerman, D., Tolberg, W., & Floyd, D. (1957) - Reaction of Polyamide Resins and Epoxy Resins. Industrial & Engineering Chemistry, 49 (7), 1091-1094Document4 pagesPeerman, D., Tolberg, W., & Floyd, D. (1957) - Reaction of Polyamide Resins and Epoxy Resins. Industrial & Engineering Chemistry, 49 (7), 1091-1094Érica RodriguesNo ratings yet
- Caballero EstevezDocument13 pagesCaballero EstevezFredy Colpas CastilloNo ratings yet
- Article MorganDocument15 pagesArticle MorganAliNo ratings yet
- 03 Ib Chemistry (SL+HL) - S1.5 Ideal GasesDocument33 pages03 Ib Chemistry (SL+HL) - S1.5 Ideal GasesricardochavezrNo ratings yet
- S5 Chemistry End of July Test Exams 2021.Document5 pagesS5 Chemistry End of July Test Exams 2021.kimberlycarls johnNo ratings yet
- DIN EN ISO 9142 - 2004-05 Bonding TestsDocument27 pagesDIN EN ISO 9142 - 2004-05 Bonding TestsAvatamanitei ConstantinNo ratings yet
- Jurnal Fistum 4Document6 pagesJurnal Fistum 4samhafiNo ratings yet
- Erosion Control / Re-Vegetation Mesh GREENAX: Technical Data SheetDocument1 pageErosion Control / Re-Vegetation Mesh GREENAX: Technical Data SheetHassan ZAFFA CISNEROSNo ratings yet
- LV Power Cables CatalogueDocument31 pagesLV Power Cables CataloguePrabhu UdayarNo ratings yet
- 40690-163685-1-PBDocument7 pages40690-163685-1-PBndafikrinNo ratings yet
- HOL Melting Point Lab ReportDocument32 pagesHOL Melting Point Lab ReportKyle Warfield100% (8)
- University of Huddersfield Repository: Original CitationDocument282 pagesUniversity of Huddersfield Repository: Original Citationdocs docsNo ratings yet
- Organometallic Chemistry OverviewDocument40 pagesOrganometallic Chemistry OverviewRabiul IslamNo ratings yet
- Raman Spectroscopy of Fe-Ti-Cr-oxides, Case Study Martian Meteorite EETA79001Document16 pagesRaman Spectroscopy of Fe-Ti-Cr-oxides, Case Study Martian Meteorite EETA79001César LópezNo ratings yet
- Document 5Document2 pagesDocument 5Dee DuanaNo ratings yet
- KP200Document4 pagesKP200DrGanesh Kame100% (1)
- Cor 199 Eng DM 160101Document2 pagesCor 199 Eng DM 160101Fraz AhmadNo ratings yet
- 03.02.2021 GCSA Revised Prices Due To DutyDocument2 pages03.02.2021 GCSA Revised Prices Due To DutyhillaireNo ratings yet
- Beckmann Rearrangement Cyclohexanone Oxime and Its Rearrangement to Ε-CaprolactamDocument4 pagesBeckmann Rearrangement Cyclohexanone Oxime and Its Rearrangement to Ε-CaprolactamElif YeşilyaprakNo ratings yet
- CHM 135H: Chemistry: Physical Principles Course Syllabus: Fall 2021 I ContactsDocument8 pagesCHM 135H: Chemistry: Physical Principles Course Syllabus: Fall 2021 I ContactsEithar FadlNo ratings yet
- Cambridge IGCSE: BIOLOGY 0610/61Document12 pagesCambridge IGCSE: BIOLOGY 0610/61Syed Maroof alamNo ratings yet
- Arenes / Benzene Chemistry: Question PaperDocument9 pagesArenes / Benzene Chemistry: Question Papervintu pvNo ratings yet
- EGE Flow SensorsDocument104 pagesEGE Flow SensorsEddboi NtelekoaNo ratings yet
- Organic Chemistry Chapter 4Document58 pagesOrganic Chemistry Chapter 4Laiba KhanNo ratings yet
- Two-Component Polysulphide Sealant Technical Data SheetDocument2 pagesTwo-Component Polysulphide Sealant Technical Data Sheetali alimNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)