You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Ed Morales - The Latin Beat - The Rhythms and Roots of Latin Music From Bossa Nova To Salsa and Beyond-Da Capo Press (2003)Document404 pagesEd Morales - The Latin Beat - The Rhythms and Roots of Latin Music From Bossa Nova To Salsa and Beyond-Da Capo Press (2003)Donald100% (6)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- MIG140 MIG170: Owner's Manual & Safety InstructionsDocument32 pagesMIG140 MIG170: Owner's Manual & Safety InstructionsPedro Luis Guaita OsorioNo ratings yet
- 10.1136@jitc 2020 001662Document8 pages10.1136@jitc 2020 001662DonaldNo ratings yet
- H.W. Siemens - Agonal Communities of Taste Law and Community in Nietzsche's Philosophy of Transvaluation (Article)Document31 pagesH.W. Siemens - Agonal Communities of Taste Law and Community in Nietzsche's Philosophy of Transvaluation (Article)DonaldNo ratings yet
- Paul Van Tongeren - Nietzsche's Greek Measure (2002) (Article)Document21 pagesPaul Van Tongeren - Nietzsche's Greek Measure (2002) (Article)DonaldNo ratings yet
- Gerasimos Xenophon Santas - The Socratic Fallacy (Article)Document16 pagesGerasimos Xenophon Santas - The Socratic Fallacy (Article)DonaldNo ratings yet
- Stephen Yablo - Go Figure - A Path Through Fictionalism (Article)Document41 pagesStephen Yablo - Go Figure - A Path Through Fictionalism (Article)DonaldNo ratings yet
- SONOR Drum Catalog 2008 WebDocument40 pagesSONOR Drum Catalog 2008 WebDonaldNo ratings yet
- Robin Waterfield - Protagoras and The Challenge of Relativism Plato's Subtlest Enemy. by Ugo Zilioli (Article)Document2 pagesRobin Waterfield - Protagoras and The Challenge of Relativism Plato's Subtlest Enemy. by Ugo Zilioli (Article)DonaldNo ratings yet
- Recent Advances in The Management of LiposarcomaDocument15 pagesRecent Advances in The Management of LiposarcomaBudhiNo ratings yet
- Breast Cancer Diagnosed During PregnancyDocument3 pagesBreast Cancer Diagnosed During PregnancyJonny CardenasNo ratings yet
- Effects of Target Volume Coverage in Superficial Medial and Lateral Breast Tissue Using Flattening Filter-Free BeamsDocument10 pagesEffects of Target Volume Coverage in Superficial Medial and Lateral Breast Tissue Using Flattening Filter-Free Beamsapi-602488644No ratings yet
- Papillary Lesions of The Breast: MRI, Ultrasound, and Mammographic AppearancesDocument8 pagesPapillary Lesions of The Breast: MRI, Ultrasound, and Mammographic Appearanceslina huraibyNo ratings yet
- Prostate CancerDocument58 pagesProstate CancerIsaac MwangiNo ratings yet
- Overal Survival of Filipinos With Head and Neck SCCADocument6 pagesOveral Survival of Filipinos With Head and Neck SCCAPrincess B. MaristelaNo ratings yet
- The Breast: Kristoff Armand E. Tan Hannah Lois Kangleon-TanDocument30 pagesThe Breast: Kristoff Armand E. Tan Hannah Lois Kangleon-TanKenneth NuñezNo ratings yet
- Bone PDFDocument14 pagesBone PDFVon HippoNo ratings yet
- The Story of An Eternal - Angelina Jolie - Abhishek ChaturvediDocument2 pagesThe Story of An Eternal - Angelina Jolie - Abhishek ChaturvediPalash SarowareNo ratings yet
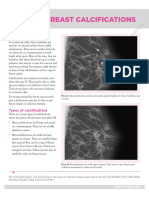
- Breast CalcificationsDocument2 pagesBreast CalcificationsAndreea MorgovanNo ratings yet
- Korelasi Aktivitas Fisik Dan Screen Time Selama Pandemi Covid-19 Pada Mahasiswa Fakultas Kedokteran Universitas Islam Bandung Tahun Akademik 2020 2021Document7 pagesKorelasi Aktivitas Fisik Dan Screen Time Selama Pandemi Covid-19 Pada Mahasiswa Fakultas Kedokteran Universitas Islam Bandung Tahun Akademik 2020 2021Najla NashirahNo ratings yet
- Cutaneous - Melanoma-Spanish Hasta 183Document233 pagesCutaneous - Melanoma-Spanish Hasta 183Ely SalazarNo ratings yet
- Solitary Thyroid NoduleDocument24 pagesSolitary Thyroid Nodulenontapat paesarochNo ratings yet
- Pancreatic Ductal Adenocarcinoma and Its Variants: Pearls and PerilsDocument20 pagesPancreatic Ductal Adenocarcinoma and Its Variants: Pearls and PerilsGeorge MogaNo ratings yet
- AG Nutrition ETHIOPIA PRESENTATION 2023 APRIDocument41 pagesAG Nutrition ETHIOPIA PRESENTATION 2023 APRIKhalid dahir aliNo ratings yet
- Jurnal Metastasis Paru 2Document25 pagesJurnal Metastasis Paru 2widiastrikNo ratings yet
- Community Assessment DamascusDocument24 pagesCommunity Assessment DamascusldhendersonNo ratings yet
- OF BLADDER CANCER" For M.Ch. Urology, Session 2020-2023Document24 pagesOF BLADDER CANCER" For M.Ch. Urology, Session 2020-2023flammator100% (1)
- Distribution and Density of Tertiary Lymphoid Structures Predict Clinical Outcome in Intrahepatic CholangiocarcinomaDocument12 pagesDistribution and Density of Tertiary Lymphoid Structures Predict Clinical Outcome in Intrahepatic Cholangiocarcinoma马三强No ratings yet
- An Introduction To Cancer Biology: When Good Cells Go BadDocument68 pagesAn Introduction To Cancer Biology: When Good Cells Go BadSavitaNo ratings yet
- Renal CancerDocument11 pagesRenal CancervishnuNo ratings yet
- Breast Lump RemovalDocument3 pagesBreast Lump RemovalMissDyYournurseNo ratings yet
- An Introduction To The 2019 ASCCP Risk Based.1Document3 pagesAn Introduction To The 2019 ASCCP Risk Based.1Elena Silvia SoldeanuNo ratings yet
- Jurnal 2018 Cervical Squamous NeoplasiaDocument78 pagesJurnal 2018 Cervical Squamous Neoplasiayeni rahmawatiNo ratings yet
- November 2020Document64 pagesNovember 2020Eric Santiago100% (1)
- Poster Laura BUCURESTIDocument1 pagePoster Laura BUCURESTIRoxana BaranNo ratings yet
- Case PresentationDocument35 pagesCase PresentationGenio AladdinNo ratings yet
- CWT Lower Gi 0Document1 pageCWT Lower Gi 0Velibor VasovicNo ratings yet
- Johnson Jerry Alan Chinese Medical Qigong Therapy Vol 5-261-280Document20 pagesJohnson Jerry Alan Chinese Medical Qigong Therapy Vol 5-261-280toanbauNo ratings yet