You might also like
- Articulo 6 Ip-10okDocument11 pagesArticulo 6 Ip-10okoscarbio2009No ratings yet
- Wilcock 2004Document6 pagesWilcock 2004everton.monteiroNo ratings yet
- Increased Threshold For TCR-Mediated Signaling Controls Self Reactivity of Intraepithelial LymphocytesDocument7 pagesIncreased Threshold For TCR-Mediated Signaling Controls Self Reactivity of Intraepithelial LymphocytesBiosynthesisNo ratings yet
- Staphylococcus Aureus Downregulates IP-10 Production and Prevents Th1 Cell Recruitment 2017Document11 pagesStaphylococcus Aureus Downregulates IP-10 Production and Prevents Th1 Cell Recruitment 2017oscarbio2009No ratings yet
- Daley JMDocument9 pagesDaley JMAnditha Namira RSNo ratings yet
- 09 J Immunol 2010 Silverman 5011 22 LeishmaniaDocument13 pages09 J Immunol 2010 Silverman 5011 22 LeishmanialilianapradaNo ratings yet
- 2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Document9 pages2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Pablo Tapia BatallaNo ratings yet
- 1659Document6 pages1659Shital BishtNo ratings yet
- Reduction of Intestinal Mucosal Immune Function in Heat Stressed Rats and Bacterial TranslocationDocument11 pagesReduction of Intestinal Mucosal Immune Function in Heat Stressed Rats and Bacterial TranslocationOwonikoko MathewNo ratings yet
- Suppression of Allergic Diarrhea in MurineDocument8 pagesSuppression of Allergic Diarrhea in MurineMeldaNo ratings yet
- Heterogeneous T Cell Responses To Beta-Lactam-Modified Self-Structures Are Observed in Penicillin-Allergic IndividualsDocument10 pagesHeterogeneous T Cell Responses To Beta-Lactam-Modified Self-Structures Are Observed in Penicillin-Allergic IndividualsEmail KosongNo ratings yet
- Ethacridinlactat Reinhardt Wounds 2005Document10 pagesEthacridinlactat Reinhardt Wounds 2005Esti SaraswatiNo ratings yet
- 1 s2.0 S0264410X11002350 MainDocument9 pages1 s2.0 S0264410X11002350 MainDidier MauricioNo ratings yet
- 1 s2.0 S1756464613002156 MainDocument9 pages1 s2.0 S1756464613002156 MainA'in Qurrota A'yuninNo ratings yet
- J Infect Dis.-2000-Hang-1738-48Document11 pagesJ Infect Dis.-2000-Hang-1738-48Anonymous hXd6PJlNo ratings yet
- Dexamethasone inhibits IL-9 production by human T cellsDocument9 pagesDexamethasone inhibits IL-9 production by human T cellsciara babyNo ratings yet
- Gangliosides Trigger Inflammatory Responses Via TLR4 in Brain GliaDocument12 pagesGangliosides Trigger Inflammatory Responses Via TLR4 in Brain GliaCaroline GravesNo ratings yet
- A Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesDocument19 pagesA Xenogenic Immune Response Towards Stz-Rinm5f Cells Reveals Cytokeratin18 As A Novel Immunogenic Antigen That May Predispose Towards Type-1 DiabetesIJAR JOURNALNo ratings yet
- MainDocument12 pagesMainMachlery KunNo ratings yet
- Monophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andDocument14 pagesMonophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andLeandro CarreñoNo ratings yet
- StatistikDocument7 pagesStatistikAkhmad Ikhwan BaidlowiNo ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsFrom EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNo ratings yet
- Hasan 2015Document9 pagesHasan 2015KianaNo ratings yet
- Stress Hyperglycemia and Innate Immune System CellsDocument9 pagesStress Hyperglycemia and Innate Immune System CellsJuan Fernando BolivarNo ratings yet
- Veterinary Immunology and ImmunopathologyDocument6 pagesVeterinary Immunology and ImmunopathologySheikh FirdousNo ratings yet
- Hyperinsulin Dan MakrofagDocument10 pagesHyperinsulin Dan Makrofageki_herawatiNo ratings yet
- Parameters to evaluate the immunostimulant effect of Zymosan A in salmonidsDocument8 pagesParameters to evaluate the immunostimulant effect of Zymosan A in salmonidsThảo ThảoNo ratings yet
- Basic Fibroblast Growth Factor Delivery Enhances ADocument11 pagesBasic Fibroblast Growth Factor Delivery Enhances AlithaNo ratings yet
- Morphological Changes in Blood Cells in A Rat Model of Heatstroke: A Pilot StudyDocument13 pagesMorphological Changes in Blood Cells in A Rat Model of Heatstroke: A Pilot StudyDaniel Roberto Sáenz GarcíaNo ratings yet
- Alteration of Intestinal Intraepithelial Lymphocytes and IncreasedDocument9 pagesAlteration of Intestinal Intraepithelial Lymphocytes and IncreaseddonkeyendutNo ratings yet
- COX-2 Expression in Neutrophils Treated Cassava LeDocument7 pagesCOX-2 Expression in Neutrophils Treated Cassava LeRena NormasariNo ratings yet
- Abstract PDFDocument36 pagesAbstract PDFDhini Kartika SariNo ratings yet
- CytokineDocument9 pagesCytokinetharaNo ratings yet
- IMM 265 - Galley Proof - 5may21 - VelikkakamDocument13 pagesIMM 265 - Galley Proof - 5may21 - VelikkakamSoraya Torres GazeNo ratings yet
- Yan Et Al (2008) - DNA Microarray Analysis of Fluconazole Resistance in A Laboratory CandidaDocument13 pagesYan Et Al (2008) - DNA Microarray Analysis of Fluconazole Resistance in A Laboratory CandidaDarfirizan SeprikaNo ratings yet
- Mass Spectrometric Analysis of The Secretome of Candida AlbicansDocument12 pagesMass Spectrometric Analysis of The Secretome of Candida AlbicansPedro ItaloNo ratings yet
- 2015 Article 700Document11 pages2015 Article 700Andreea KNo ratings yet
- Mice, A Model of Human Chronic / Nonresolving Inflammation in gp91Document9 pagesMice, A Model of Human Chronic / Nonresolving Inflammation in gp91Saurav KumarNo ratings yet
- Identification and Characterization of A Human IL-10 Receptor AntagonistDocument4 pagesIdentification and Characterization of A Human IL-10 Receptor AntagonistMathieu VigneauNo ratings yet
- Animals 11 01286Document13 pagesAnimals 11 01286Alejandro OviedoNo ratings yet
- Tian 2016Document13 pagesTian 2016Epiphany SonderNo ratings yet
- Pubmed5Document11 pagesPubmed5Febby AuliaNo ratings yet
- J Biol Chem - 2004-Beller-46129-34Document7 pagesJ Biol Chem - 2004-Beller-46129-34api-212391856No ratings yet
- Investigation of Energy Metabolic Dynamism in Hyperthermia Resistant Ovarian and Uterine Cancer Cells Under Heat StressDocument11 pagesInvestigation of Energy Metabolic Dynamism in Hyperthermia Resistant Ovarian and Uterine Cancer Cells Under Heat Stress29 - Heaven JosiahNo ratings yet
- КАПСУЛИРУЕМСЯ, БРАТВАDocument33 pagesКАПСУЛИРУЕМСЯ, БРАТВААлексей КочетовNo ratings yet
- Recombinant Lactobacillus Plantarum Inhibits House Dust Mite-Specific T-Cell ResponsesDocument7 pagesRecombinant Lactobacillus Plantarum Inhibits House Dust Mite-Specific T-Cell ResponsesIdelsy GradailleNo ratings yet
- Alloreactivity Instructed by Invariant Human Invariant NKT Cells DisplayDocument10 pagesAlloreactivity Instructed by Invariant Human Invariant NKT Cells DisplayLeandro CarreñoNo ratings yet
- Art 3A10.1186 2F1476 9255 5 16 - 2Document11 pagesArt 3A10.1186 2F1476 9255 5 16 - 2Lilis RohaetiNo ratings yet
- Growth Arrest Specific Gene 2 in Tilapia (Oreochromis Niloticus) : Molecular Characterization and Functional Analysis Under Low-Temperature StressDocument11 pagesGrowth Arrest Specific Gene 2 in Tilapia (Oreochromis Niloticus) : Molecular Characterization and Functional Analysis Under Low-Temperature Stressbing masturNo ratings yet
- Structure Kestose and NystoseDocument6 pagesStructure Kestose and NystoseJavier MendozaNo ratings yet
- Department of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University, Columbus, Ohio 43210-1093Document9 pagesDepartment of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University, Columbus, Ohio 43210-1093FrontiersNo ratings yet
- Microorganisms 08 01175Document14 pagesMicroorganisms 08 01175Karima El-yagoubiNo ratings yet
- Immunological: Section of LaryngologyDocument3 pagesImmunological: Section of LaryngologymfhfhfNo ratings yet
- BMC ImmunologyDocument9 pagesBMC ImmunologyYuniarNo ratings yet
- J Immunol 2007 Rydström 5789 801Document14 pagesJ Immunol 2007 Rydström 5789 801susana1616No ratings yet
- 10.1002@1097 0290 (20001020) 70 2 217 Aid Bit11 3.0.co 2 WDocument8 pages10.1002@1097 0290 (20001020) 70 2 217 Aid Bit11 3.0.co 2 WVenkata Suryanarayana GorleNo ratings yet
- Rendahnya Vitd3Document17 pagesRendahnya Vitd3ervina 119260026No ratings yet
- Loss of The Interleukin-6 Receptor Causes Immunodeficiency, Atopy, and Abnormal Inflammatory ResponsesDocument13 pagesLoss of The Interleukin-6 Receptor Causes Immunodeficiency, Atopy, and Abnormal Inflammatory ResponsesSyed Irfan RazaNo ratings yet
- Patch Testing of Experimentally Sensitized Beagle Dogs - Development of A Model For Skin Lesions of Atopic Dermatitis (Pages 95-102)Document8 pagesPatch Testing of Experimentally Sensitized Beagle Dogs - Development of A Model For Skin Lesions of Atopic Dermatitis (Pages 95-102)jenNo ratings yet
- Immunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988From EverandImmunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988J. F. BachNo ratings yet
- Quality Control ResultsDocument4 pagesQuality Control ResultsVan RivNo ratings yet
- 2005, Storni, Immunity in Response To Ag DeliveryDocument23 pages2005, Storni, Immunity in Response To Ag DeliveryVan RivNo ratings yet
- 1997, Hanes, New Advances in MicrosphereDocument23 pages1997, Hanes, New Advances in MicrosphereVan RivNo ratings yet
- 1995, Hanes, Polymer Microspheres VaccineDocument24 pages1995, Hanes, Polymer Microspheres VaccineVan RivNo ratings yet
- Early Human Development: A. Nicolai, M. Ferrara, C. Schiavariello, F. Gentile, M.E. Grande, C. Alessandroni, F. MidullaDocument5 pagesEarly Human Development: A. Nicolai, M. Ferrara, C. Schiavariello, F. Gentile, M.E. Grande, C. Alessandroni, F. MidullalalaNo ratings yet
- Final Presentation (Stock Pitch) - ModernaDocument8 pagesFinal Presentation (Stock Pitch) - ModernaAarushiNo ratings yet
- Tuberculous MeningitisDocument8 pagesTuberculous MeningitisEchizen RukawaNo ratings yet
- Management of Erythema Multiforme Secondary To Herpes Simplex by Systemic Acyclovir and Topical Corticosteroid A Case Report 45 49 PDFDocument5 pagesManagement of Erythema Multiforme Secondary To Herpes Simplex by Systemic Acyclovir and Topical Corticosteroid A Case Report 45 49 PDFindrayaniNo ratings yet
- Hiv & Aids Awareness ProgramDocument8 pagesHiv & Aids Awareness ProgramKenzie WalshNo ratings yet
- Impact of Blood Group in Dengue: A Study: Original Research ArticleDocument5 pagesImpact of Blood Group in Dengue: A Study: Original Research ArticleMuhammad AfzaalNo ratings yet
- Opportunistic Mycoses: Portal of Entry: Respiratory, Mucocutaneous Infectious Particles: Conidia, MyceliaDocument4 pagesOpportunistic Mycoses: Portal of Entry: Respiratory, Mucocutaneous Infectious Particles: Conidia, MyceliaPinay YaunNo ratings yet
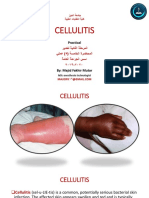
- Cellulitis Infection Causes, Symptoms and TreatmentDocument11 pagesCellulitis Infection Causes, Symptoms and TreatmentAli dhyaaNo ratings yet
- ChorioretinitisDocument19 pagesChorioretinitisilmansyahandiNo ratings yet
- AllergyDocument26 pagesAllergyPuskesmas Kabe0% (1)
- Treg CellsDocument15 pagesTreg Cellsmr.hakku.jetaNo ratings yet
- BNS-102 Applied Science Exam for Post Basic Nursing StudentsDocument6 pagesBNS-102 Applied Science Exam for Post Basic Nursing StudentsManu CvNo ratings yet
- Eur J Immunol - 2020 - Hanitsch - Treatment and Management of Primary Antibody Deficiency German InterdisciplinaryDocument15 pagesEur J Immunol - 2020 - Hanitsch - Treatment and Management of Primary Antibody Deficiency German InterdisciplinaryRSU Trijaya medical centerNo ratings yet
- (醫學筆記) 抗生素使用 Antibiotics part.1 概論+β-lactam: by kphsien Published 97 CommentsDocument8 pages(醫學筆記) 抗生素使用 Antibiotics part.1 概論+β-lactam: by kphsien Published 97 Comments蔡宏達No ratings yet
- What Is Neisseria GonorrhoeaeDocument8 pagesWhat Is Neisseria GonorrhoeaeRosNo ratings yet
- 24.18.00 - Chikungunya2 DR RAGADocument48 pages24.18.00 - Chikungunya2 DR RAGAjcvh24No ratings yet
- Pneumonia PediatricsDocument11 pagesPneumonia PediatricsJakobus Benny SalimNo ratings yet
- მეზენქიმური ღეროვანი უჯრედებიDocument1 pageმეზენქიმური ღეროვანი უჯრედებიEMD GROUPNo ratings yet
- Presentation Outline Group: Blood Type Leader: Ratna Ningsih Members: Fahrul RojiDocument5 pagesPresentation Outline Group: Blood Type Leader: Ratna Ningsih Members: Fahrul RojiFahrul RoziNo ratings yet
- Risk For InfectionDocument2 pagesRisk For InfectionALEKS MONTECINO JIMENEZNo ratings yet
- Table of blood group systems v. 9.0Document3 pagesTable of blood group systems v. 9.0AnthonyNo ratings yet
- Inflammation: InfectionDocument11 pagesInflammation: InfectionRenBautistaNo ratings yet
- Lasercyte DX Dot Plot Poster enDocument2 pagesLasercyte DX Dot Plot Poster enSpital Veterinar AndivetNo ratings yet
- Gram Positive & Gram Negative Cocci Infections (Staphylococcus, Streptococcus, NeisseriaDocument33 pagesGram Positive & Gram Negative Cocci Infections (Staphylococcus, Streptococcus, NeisseriaRebeka Costantina Weriditi0% (1)
- Vaccine Safety HandbookDocument40 pagesVaccine Safety HandbookleostraussNo ratings yet
- Day 37 - Daily MCQ Workout - 40 Revision MCQsDocument5 pagesDay 37 - Daily MCQ Workout - 40 Revision MCQsBobbyNo ratings yet
- Basic - Haematology Notes - Updated AnggelosDocument123 pagesBasic - Haematology Notes - Updated AnggelosLorenz SmallNo ratings yet
- Infection Control, Safety First Aid and Personal WellnessDocument9 pagesInfection Control, Safety First Aid and Personal WellnessJacinta MalamionNo ratings yet
- Autoimmune Hepatitis HandoutDocument2 pagesAutoimmune Hepatitis HandoutClarisse SantosNo ratings yet
- ReviewDocument13 pagesReviewChannelle Venice UnidadNo ratings yet