You might also like
- tmp490 TMPDocument11 pagestmp490 TMPFrontiersNo ratings yet
- Taler 2013Document5 pagesTaler 2013ArouetNo ratings yet
- Knockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3Document12 pagesKnockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3郭耿良No ratings yet
- TMP 7 DD1Document12 pagesTMP 7 DD1FrontiersNo ratings yet
- Biocompatibility Evaluation of Tramadol Loaded Nanoparticulate SystemsDocument2 pagesBiocompatibility Evaluation of Tramadol Loaded Nanoparticulate SystemsVeronica NedelcuNo ratings yet
- Karen 2013 Effect of Propofol in The Immature Rat Brain On Short - and Long-Term Neurodevelopmental OutcomeDocument11 pagesKaren 2013 Effect of Propofol in The Immature Rat Brain On Short - and Long-Term Neurodevelopmental OutcomePablo MartinezNo ratings yet
- Neurplast en El Estres y ATDduman1999 PDFDocument11 pagesNeurplast en El Estres y ATDduman1999 PDFmarielaNo ratings yet
- Naskar Et Al-2013-Journal of Pineal ResearchDocument9 pagesNaskar Et Al-2013-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Content ServerpaperDocument8 pagesContent ServerpaperLuis Ignacio Baeza GonzálezNo ratings yet
- Elaine 20-10 Cellular and Molecular Basis For Stress-Induced Depressionmp2016118aDocument8 pagesElaine 20-10 Cellular and Molecular Basis For Stress-Induced Depressionmp2016118aElaine BedinNo ratings yet
- Welter2008Document6 pagesWelter2008RafaelNo ratings yet
- Abstracts For IHC WebsiteDocument193 pagesAbstracts For IHC WebsitebradbeloteNo ratings yet
- The Effects of Repeated Restraint Stress On The Synaptic Plasticity in The Inner Molecular Layer of Mouse Dentate GyrusDocument13 pagesThe Effects of Repeated Restraint Stress On The Synaptic Plasticity in The Inner Molecular Layer of Mouse Dentate Gyrussonjeonggyu87No ratings yet
- Symposium 2023 AbstractsDocument21 pagesSymposium 2023 Abstractsmail2umamaheswari15No ratings yet
- Murphy 1990Document5 pagesMurphy 1990TANSUNo ratings yet
- Ramirez 2015Document17 pagesRamirez 2015jarecot100% (1)
- A preliminary study of the effect of PPAR- γ agonist from NuSE on the biogenesis of rat's infants brain mitochondria & D1 dopamine receptorDocument4 pagesA preliminary study of the effect of PPAR- γ agonist from NuSE on the biogenesis of rat's infants brain mitochondria & D1 dopamine receptorMS Clinic Anti aging & AestheticNo ratings yet
- Progesterone Synthesis in The Nervous System: Implications For Myelination and Myelin RepairDocument22 pagesProgesterone Synthesis in The Nervous System: Implications For Myelination and Myelin Repair147Anastaysia Anisa HaribaikNo ratings yet
- Fan2016 Tissue Repair and Regeneration PDFDocument15 pagesFan2016 Tissue Repair and Regeneration PDFDavid LuNo ratings yet
- TMP 3 BD8Document12 pagesTMP 3 BD8FrontiersNo ratings yet
- Melatonina (2006) Cheung R, Safety IV RatasDocument7 pagesMelatonina (2006) Cheung R, Safety IV RatasAlumno del Doctorado FarmacologiaNo ratings yet
- KrsmanovickDocument7 pagesKrsmanovickKayro Lopez RamírezNo ratings yet
- Journal of Neurochemistry Doi: 10.1111/j.1471-4159.2009.06433.xDocument16 pagesJournal of Neurochemistry Doi: 10.1111/j.1471-4159.2009.06433.xJulie Susan SamNo ratings yet
- 10 1016@j Euroneuro 2018 11 841Document2 pages10 1016@j Euroneuro 2018 11 841gabrielledireitoNo ratings yet
- Valproic Acid, But Not Lamotrigine, Suppresses Seizure-Induced C-Fos and C-Jun mRNA ExpressionDocument5 pagesValproic Acid, But Not Lamotrigine, Suppresses Seizure-Induced C-Fos and C-Jun mRNA ExpressionFrontiersNo ratings yet
- Goldstein 1996 Role of The Amygdala in The Coordination of BehavioralDocument12 pagesGoldstein 1996 Role of The Amygdala in The Coordination of BehavioralDayna DamianiNo ratings yet
- Iranian Journal of Basic Medical SciencesDocument10 pagesIranian Journal of Basic Medical SciencesmusmuafeerahNo ratings yet
- Correcting Mir92A-Vgat-Mediated Gabaergic Dysfunctions Rescues Human Tau-Induced Anxiety in MiceDocument13 pagesCorrecting Mir92A-Vgat-Mediated Gabaergic Dysfunctions Rescues Human Tau-Induced Anxiety in MicegabrielledireitoNo ratings yet
- Gelatinase Activity Imaged by Activatable Cell-Penetrating Peptides in Cell-Based and in Vivo Models of StrokeDocument13 pagesGelatinase Activity Imaged by Activatable Cell-Penetrating Peptides in Cell-Based and in Vivo Models of StrokeKeyvan AmiriNo ratings yet
- Neurology & NeurophysiologyDocument6 pagesNeurology & NeurophysiologyRakesh KumarNo ratings yet
- Ovarian Hormones Ameliorate Memory Impairment, Cholinergic Deficit, Neuronal Apoptosis and Astrogliosis in A Rat Model of Alzheimer's DiseaseDocument9 pagesOvarian Hormones Ameliorate Memory Impairment, Cholinergic Deficit, Neuronal Apoptosis and Astrogliosis in A Rat Model of Alzheimer's DiseaseHümay ÜnalNo ratings yet
- Pinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.Document8 pagesPinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.scribdNo ratings yet
- The Nucleus Reuniens: A Key Node in The Neurocircuitry of Stress and DepressionDocument8 pagesThe Nucleus Reuniens: A Key Node in The Neurocircuitry of Stress and DepressionEszter SzerencsiNo ratings yet
- Refractive and CataractDocument61 pagesRefractive and CataractClarissaL12No ratings yet
- AsenapineTransdermalPatchVsPlaceboPosterAbstract CITROME BiolPsych2019Document1 pageAsenapineTransdermalPatchVsPlaceboPosterAbstract CITROME BiolPsych2019Leslie CitromeNo ratings yet
- Chronic Stress Changes Prepulse Inhibition After Amphetamine Challenge: The Role of The Dopaminergic SystemDocument13 pagesChronic Stress Changes Prepulse Inhibition After Amphetamine Challenge: The Role of The Dopaminergic SystemArlette CárdenasNo ratings yet
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenNo ratings yet
- Toxicidade Da Avermectina em Bovinos Com Menos de Trinta Dias de IdadeDocument10 pagesToxicidade Da Avermectina em Bovinos Com Menos de Trinta Dias de IdadeErick AlvesNo ratings yet
- Ultra-High-Dose Methylcobalamin in ALS - A Long Term Phase II-III Randomised Controlled StudyDocument7 pagesUltra-High-Dose Methylcobalamin in ALS - A Long Term Phase II-III Randomised Controlled StudyAurelia Sartika TongkuNo ratings yet
- MitochondrialDocument13 pagesMitochondrialAndrea CamachoNo ratings yet
- Pharmacological Manipulation of Serotonin Receptors During Brain 2017Document4 pagesPharmacological Manipulation of Serotonin Receptors During Brain 2017Marco GiammancoNo ratings yet
- Evaluation of Antidepressant Potential of Agmatine in Diabetes Induce Depression in RatsDocument6 pagesEvaluation of Antidepressant Potential of Agmatine in Diabetes Induce Depression in RatsnikuNo ratings yet
- J Jns 2004 12 011Document10 pagesJ Jns 2004 12 011Viviana LunaNo ratings yet
- PpokDocument7 pagesPpokSkoy KuyNo ratings yet
- 33 Pejon Et Al., 2022 IJSMDocument11 pages33 Pejon Et Al., 2022 IJSMLucianaNo ratings yet
- Matrix MetalloproteinaseDocument4 pagesMatrix Metalloproteinasetest2012No ratings yet
- Ijpbs 2011Document8 pagesIjpbs 2011Ramakrishna DevakiNo ratings yet
- Epigallocatechin-3-Gallate Suppresses TNF-of MMP-1 and - 3 in Rheumatoid Arthritis Synovial WbroblastsDocument7 pagesEpigallocatechin-3-Gallate Suppresses TNF-of MMP-1 and - 3 in Rheumatoid Arthritis Synovial WbroblastsTharuka ChandrasiriNo ratings yet
- Inhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Document13 pagesInhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Apróné Török IbolyaNo ratings yet
- Synaptic Expression of Glutamate Receptor After Encoding of Fear Memory in The Rat AmygdalaDocument38 pagesSynaptic Expression of Glutamate Receptor After Encoding of Fear Memory in The Rat AmygdalaGia KuteliaNo ratings yet
- Maternal Care and Hipoc Weaver and Meaney PDFDocument6 pagesMaternal Care and Hipoc Weaver and Meaney PDFmarielaNo ratings yet
- Arora 2009Document6 pagesArora 2009ece142No ratings yet
- Neurobiology of Learning and MemoryDocument7 pagesNeurobiology of Learning and MemoryNothingNo ratings yet
- Ramos Et Al 2007 DefinitivoDocument9 pagesRamos Et Al 2007 Definitivolucasmonje1978No ratings yet
- Effects of Melatonin On Human Mood and Performance: Research ReportsDocument7 pagesEffects of Melatonin On Human Mood and Performance: Research ReportsSriram BalajiNo ratings yet
- Sub-Chronic Nandrolone Treatment Modifies Neurochemical and Behavioral Effects of Amphetamine and 3,4-Methylenedioxymethamphetamine (MDMA) in RatsDocument11 pagesSub-Chronic Nandrolone Treatment Modifies Neurochemical and Behavioral Effects of Amphetamine and 3,4-Methylenedioxymethamphetamine (MDMA) in RatsSanaNo ratings yet
- Antiepileptic Drugs Under Clinical Trials (Amit Mehta, 1911856, 7th Sem) 1Document21 pagesAntiepileptic Drugs Under Clinical Trials (Amit Mehta, 1911856, 7th Sem) 1vishal sharmaNo ratings yet
- Restorative Effects of Stimulating Medullary Raphe After Spinal Cord InjuryDocument14 pagesRestorative Effects of Stimulating Medullary Raphe After Spinal Cord Injurycamillemae24No ratings yet
- Journal 2Document9 pagesJournal 2Suryanti SultanNo ratings yet
- Liu Et Al-2013-Journal of Pineal ResearchDocument10 pagesLiu Et Al-2013-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Naskar Et Al-2013-Journal of Pineal ResearchDocument9 pagesNaskar Et Al-2013-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Lima-Cabello Et Al-2014-Journal of Pineal ResearchDocument9 pagesLima-Cabello Et Al-2014-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- The Microtubule Cytoskeleton Acts As A Key Downstream Effector of Neurotransmitter SignalingDocument8 pagesThe Microtubule Cytoskeleton Acts As A Key Downstream Effector of Neurotransmitter SignalingElena Martínez CelisNo ratings yet
- Kaur Et Al-2010-Journal of Pineal ResearchDocument9 pagesKaur Et Al-2010-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Lacoste Et Al-2015-Journal of Pineal ResearchDocument21 pagesLacoste Et Al-2015-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Tau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersDocument30 pagesTau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersElena Martínez CelisNo ratings yet
- G-Actina Espinas DendriticasDocument3 pagesG-Actina Espinas DendriticasElena Martínez CelisNo ratings yet
- Phosporilation Tau EsquemaDocument9 pagesPhosporilation Tau EsquemaElena Martínez CelisNo ratings yet
- MELATONINA F ACTINA Matsui - Et - Al-1997-Journal - of - Pineal - ResearchDocument7 pagesMELATONINA F ACTINA Matsui - Et - Al-1997-Journal - of - Pineal - ResearchElena Martínez CelisNo ratings yet
- Actin-Based Plasticity in Dendritic Spines: References and NotesDocument6 pagesActin-Based Plasticity in Dendritic Spines: References and NotesElena Martínez CelisNo ratings yet
- Calcium in SpinesDocument6 pagesCalcium in SpinesElena Martínez CelisNo ratings yet
- Radiographic Positioning DogDocument1 pageRadiographic Positioning DogElena Martínez Celis100% (2)
- Sakui, K., & Cowie, N. (2012) - The Dark Side of Motivation - Teachers' Perspectives On 'Unmotivation'. ELTJ, 66 (2), 205-213.Document9 pagesSakui, K., & Cowie, N. (2012) - The Dark Side of Motivation - Teachers' Perspectives On 'Unmotivation'. ELTJ, 66 (2), 205-213.Robert HutchinsonNo ratings yet
- 16783Document51 pages16783uddinnadeemNo ratings yet
- Subject Manual Tle 7-8Document11 pagesSubject Manual Tle 7-8Rhayan Dela Cruz DaquizNo ratings yet
- Miniature Daisy: Crochet Pattern & InstructionsDocument8 pagesMiniature Daisy: Crochet Pattern & Instructionscaitlyn g100% (1)
- PDFDocument10 pagesPDFerbariumNo ratings yet
- IEC TC 56 Dependability PDFDocument8 pagesIEC TC 56 Dependability PDFsaospieNo ratings yet
- World Insurance Report 2017Document36 pagesWorld Insurance Report 2017deolah06No ratings yet
- Case CapsuleDocument8 pagesCase CapsuleLiza BulsaraNo ratings yet
- B I o G R A P H yDocument17 pagesB I o G R A P H yRizqia FitriNo ratings yet
- Canoe Matlab 001Document58 pagesCanoe Matlab 001Coolboy RoadsterNo ratings yet
- 2022 Mable Parker Mclean Scholarship ApplicationDocument2 pages2022 Mable Parker Mclean Scholarship Applicationapi-444959661No ratings yet
- LSCM Course OutlineDocument13 pagesLSCM Course OutlineDeep SachetiNo ratings yet
- Antena TelnetDocument4 pagesAntena TelnetMarco PiambaNo ratings yet
- Department of Education: Republic of The PhilippinesDocument1 pageDepartment of Education: Republic of The PhilippinesJonathan CayatNo ratings yet
- Activity On Noli Me TangereDocument5 pagesActivity On Noli Me TangereKKKNo ratings yet
- EPAS 11 - Q1 - W1 - Mod1Document45 pagesEPAS 11 - Q1 - W1 - Mod1Alberto A. FugenNo ratings yet
- Durex 'S Marketing Strategy in VietnamDocument45 pagesDurex 'S Marketing Strategy in VietnamPham Nguyen KhoiNo ratings yet
- Enochian Dragon Ritual PDFDocument4 pagesEnochian Dragon Ritual PDFDenis NantelNo ratings yet
- Journal of Biology EducationDocument13 pagesJournal of Biology EducationFarah ArrumyNo ratings yet
- Bullshit System v0.5Document40 pagesBullshit System v0.5ZolaniusNo ratings yet
- Introduction To Screenwriting UEADocument12 pagesIntroduction To Screenwriting UEAMartín SalasNo ratings yet
- Rana2 Compliment As Social StrategyDocument12 pagesRana2 Compliment As Social StrategyRanaNo ratings yet
- Unsuccessful MT-SM DeliveryDocument2 pagesUnsuccessful MT-SM DeliveryPitam MaitiNo ratings yet
- BrochureDocument3 pagesBrochureapi-400730798No ratings yet
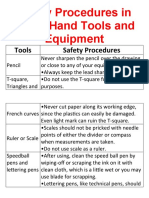
- Safety Procedures in Using Hand Tools and EquipmentDocument12 pagesSafety Procedures in Using Hand Tools and EquipmentJan IcejimenezNo ratings yet
- Wner'S Anual: Led TVDocument32 pagesWner'S Anual: Led TVErmand WindNo ratings yet
- 2022 WR Extended VersionDocument71 pages2022 WR Extended Versionpavankawade63No ratings yet
- Mechanical Production Engineer Samphhhhhle ResumeDocument2 pagesMechanical Production Engineer Samphhhhhle ResumeAnirban MazumdarNo ratings yet
- Workbook Group TheoryDocument62 pagesWorkbook Group TheoryLi NguyenNo ratings yet
- Pg2022 ResultDocument86 pagesPg2022 ResultkapilNo ratings yet