You might also like
- Neurological Physiotherapy Evaluation FormDocument13 pagesNeurological Physiotherapy Evaluation FormShruti100% (10)
- Applied Anthropology: By: DR Habibullah Abbasi Assistant Professor Center For Environmental Science, UosDocument13 pagesApplied Anthropology: By: DR Habibullah Abbasi Assistant Professor Center For Environmental Science, UosZain MemonNo ratings yet
- The Complexity of Molecular Processes in Osteoarthritis of The Knee JointDocument10 pagesThe Complexity of Molecular Processes in Osteoarthritis of The Knee JointRita DiabNo ratings yet
- Research developments in musculoskeletal disordersDocument3 pagesResearch developments in musculoskeletal disordersBrandon Felipe Tejeda GarzonNo ratings yet
- Occurrence of Tendon Pathologies in MetabolicDocument10 pagesOccurrence of Tendon Pathologies in MetabolicSebastián Jiménez GaticaNo ratings yet
- Agm2 2 147 PDFDocument10 pagesAgm2 2 147 PDFAntonioNo ratings yet
- 1 s2.0 S204908012101027X Main 1Document10 pages1 s2.0 S204908012101027X Main 1Amirah FaridahNo ratings yet
- Clinical Aspects, Pathology and Pathophysiology of OsteoarthritisDocument3 pagesClinical Aspects, Pathology and Pathophysiology of OsteoarthritisOscar NgNo ratings yet
- Antioxidants: Impacts of Green Tea On Joint and Skeletal Muscle Health: Prospects of Translational NutritionDocument28 pagesAntioxidants: Impacts of Green Tea On Joint and Skeletal Muscle Health: Prospects of Translational NutritionKarina CifuentesNo ratings yet
- Chondroprotective Effects of The Combination ChondroitinDocument9 pagesChondroprotective Effects of The Combination Chondroitincursotplo2021No ratings yet
- Abnormalities of Neuromuscular Anatomy in Diverticular DiseaseDocument5 pagesAbnormalities of Neuromuscular Anatomy in Diverticular DiseaseHasan Al BasriNo ratings yet
- Osteoarthritis: Pathology, Diagnosis, and Treatment OptionsDocument19 pagesOsteoarthritis: Pathology, Diagnosis, and Treatment OptionsTrino Rangel FonsecaNo ratings yet
- Bailey Downs2012Document13 pagesBailey Downs2012AlexandraNo ratings yet
- OsteoartritisDocument11 pagesOsteoartritisSavira Rizka MeidySNo ratings yet
- Biomechanical Factors in OsteoarthritisDocument9 pagesBiomechanical Factors in OsteoarthritisQuangTiến ĐỗNo ratings yet
- Ageing and OsteoarthritisDocument9 pagesAgeing and OsteoarthritisHalim Muhammad SatriaNo ratings yet
- Natural History: Changes in Disc Structure With Aging and DegenerationDocument11 pagesNatural History: Changes in Disc Structure With Aging and Degenerationfreska ayu wardhaniNo ratings yet
- Links Between Osteoarthritis and Type 2 DiabetesDocument22 pagesLinks Between Osteoarthritis and Type 2 DiabetesRafael EsdrasNo ratings yet
- Effects of Glucosamine and Chondroitin Sulfate On Cartilage Metabolism in OA. Outlook On Other Nutrient Partners Especially Omega-3 Fatty AcidsDocument17 pagesEffects of Glucosamine and Chondroitin Sulfate On Cartilage Metabolism in OA. Outlook On Other Nutrient Partners Especially Omega-3 Fatty Acidsharvestmoon.harvestingtimeNo ratings yet
- Treating Tendonopathy 2015Document12 pagesTreating Tendonopathy 2015Victor Andrés Olivares IbarraNo ratings yet
- Author: Section Editor: Deputy Editor:: Pathogenesis of Systemic Sclerosis (Scleroderma)Document24 pagesAuthor: Section Editor: Deputy Editor:: Pathogenesis of Systemic Sclerosis (Scleroderma)zakiNo ratings yet
- Molecules 22 00105 v2Document26 pagesMolecules 22 00105 v2Emily BittencourtNo ratings yet
- Extracellular Vesicles Derived From HumanDocument21 pagesExtracellular Vesicles Derived From HumanMartín MamchurNo ratings yet
- Review Article: Aging and Osteoarthritis: An Inevitable Encounter?Document7 pagesReview Article: Aging and Osteoarthritis: An Inevitable Encounter?Iicha Phine'ToxicNo ratings yet
- 10.1007@s10067 020 05150 ZDocument8 pages10.1007@s10067 020 05150 ZEriC. ChaN.No ratings yet
- Alergi Vs OsteoporosisDocument15 pagesAlergi Vs OsteoporosisclaudyNo ratings yet
- Mechanobiology and Diseases of Mechanotransduction: Donald E IngberDocument14 pagesMechanobiology and Diseases of Mechanotransduction: Donald E IngberTezar AndreanNo ratings yet
- Osteoarthritis Pathogenesis A Review of Molecular MechanismsDocument11 pagesOsteoarthritis Pathogenesis A Review of Molecular Mechanismsjake zapataNo ratings yet
- DeGroot Et Al. - 2004 - Accumulation of Advanced Glycation End Products As A Molecular Mechanism For Aging As A Risk Factor in OsteoarthDocument9 pagesDeGroot Et Al. - 2004 - Accumulation of Advanced Glycation End Products As A Molecular Mechanism For Aging As A Risk Factor in OsteoarthYuki SATAKENo ratings yet
- Articular Cartilage and Changes in Arthritis An Introduction: Cell Biology of OsteoarthritisDocument7 pagesArticular Cartilage and Changes in Arthritis An Introduction: Cell Biology of OsteoarthritisBasheer HamidNo ratings yet
- Loukov 2018Document9 pagesLoukov 2018Jonathan Billy ChristianNo ratings yet
- Literature Review OsteoarthritisDocument10 pagesLiterature Review Osteoarthritisafdtbbhtz100% (1)
- Adiponectin SynovialDocument12 pagesAdiponectin Synovialiosif iliaNo ratings yet
- Understanding the Pathogenesis and Pathophysiology of Rheumatoid ArthritisDocument6 pagesUnderstanding the Pathogenesis and Pathophysiology of Rheumatoid ArthritisGeorge WinchesterNo ratings yet
- Review Injeksi Intraartikular Pada OA LututDocument12 pagesReview Injeksi Intraartikular Pada OA LututUmmu Qonitah AfnidawatiNo ratings yet
- Review Article: Combating Osteoarthritis Through Stem Cell Therapies by Rejuvenating Cartilage: A ReviewDocument13 pagesReview Article: Combating Osteoarthritis Through Stem Cell Therapies by Rejuvenating Cartilage: A ReviewAlvin GfNo ratings yet
- Obesity and Osteoarthritis: What Are The Links?: Francis Berenbaum, Je Re Mie SellamDocument2 pagesObesity and Osteoarthritis: What Are The Links?: Francis Berenbaum, Je Re Mie SellamHervi LaksariNo ratings yet
- Biomolecules: TRPV Channels in Osteoarthritis: A Comprehensive ReviewDocument25 pagesBiomolecules: TRPV Channels in Osteoarthritis: A Comprehensive ReviewTedNo ratings yet
- Correlacion Osteoporosis y Enfermedad CardiovascularDocument3 pagesCorrelacion Osteoporosis y Enfermedad CardiovascularCorin Boice TelloNo ratings yet
- Clinical Review Paget's Disease of BoneDocument5 pagesClinical Review Paget's Disease of BoneIzni FatihahNo ratings yet
- Evidence and Perspectives of Cell Senescence in Neurodegenerative DiseasesDocument11 pagesEvidence and Perspectives of Cell Senescence in Neurodegenerative DiseasesKarel GuevaraNo ratings yet
- JCM 11 06013Document12 pagesJCM 11 06013ryanNo ratings yet
- Osteoarthritis Guide: Causes, Symptoms, TreatmentsDocument3 pagesOsteoarthritis Guide: Causes, Symptoms, TreatmentsFrancisco Palma CarvajalNo ratings yet
- Epigenetics and Aging: ReviewDocument20 pagesEpigenetics and Aging: ReviewDeepak PanwarNo ratings yet
- Pathogenesis of Systemic Sclerosis Scleroderma UpToDateDocument27 pagesPathogenesis of Systemic Sclerosis Scleroderma UpToDateLê Hạ Long HảiNo ratings yet
- Bone Metabolism and Fracture Risk in Type 2 Diabetes MellitusDocument12 pagesBone Metabolism and Fracture Risk in Type 2 Diabetes MellituslusianapsNo ratings yet
- Moos 2020 Klotho Pathways Myelination DisordeDocument12 pagesMoos 2020 Klotho Pathways Myelination DisordeFiterman AdrianNo ratings yet
- Pathogenesis of OADocument5 pagesPathogenesis of OAsenthilNo ratings yet
- 3 Oa RMDocument15 pages3 Oa RMAhmad Nur Fikri AbrorNo ratings yet
- The pathogenesis of tendinopathy from a molecular perspectiveDocument12 pagesThe pathogenesis of tendinopathy from a molecular perspectiveSharon BalboaNo ratings yet
- Sarcopenia Aging-Related Loss of Muscle Mass and FunctionDocument143 pagesSarcopenia Aging-Related Loss of Muscle Mass and FunctionVictoria MNo ratings yet
- Ni Hms 825760Document13 pagesNi Hms 825760Dea MustikaNo ratings yet
- Biomechanics and Pathomechanisms of Osteoarthritis: Christian Egloff, Thomas Hügle, Victor ValderrabanoDocument14 pagesBiomechanics and Pathomechanisms of Osteoarthritis: Christian Egloff, Thomas Hügle, Victor ValderrabanobertouwNo ratings yet
- Ages and Bone Ageing in Diabetes Mellitus 2155 6156.1000276Document8 pagesAges and Bone Ageing in Diabetes Mellitus 2155 6156.1000276muhammad fajrinNo ratings yet
- Amyloid Beta Multiple Mechanisms of Toxicity and Only Some Protective EffectsDocument16 pagesAmyloid Beta Multiple Mechanisms of Toxicity and Only Some Protective EffectsEvernim OmpacanNo ratings yet
- McInnes y Schett - 2017 - Pathogenetic Insights From The Treatment of RheumaDocument20 pagesMcInnes y Schett - 2017 - Pathogenetic Insights From The Treatment of Rheumabarrera2001No ratings yet
- Czajkowska 2022Document13 pagesCzajkowska 2022Omar AhmedNo ratings yet
- Killinger 2010Document8 pagesKillinger 2010Andrea TixeNo ratings yet
- 387 06 13 ArticleDocument5 pages387 06 13 ArticleREHAB AMBULATORIO HOSPITAL VITARTENo ratings yet
- 2019 Article 369 PDFDocument10 pages2019 Article 369 PDFAziir 'arNo ratings yet
- Clinical Genetics and Genomics of AgingFrom EverandClinical Genetics and Genomics of AgingJuan Carlos Gomez-VerjanNo ratings yet
- Osteoimmunology: Interactions of the Immune and Skeletal SystemsFrom EverandOsteoimmunology: Interactions of the Immune and Skeletal SystemsJoseph LorenzoNo ratings yet
- OSTEOARTHRITISDocument2 pagesOSTEOARTHRITISAssasination ClassroomNo ratings yet
- Vitamins: Functions, Deficiencies and Food SourcesDocument54 pagesVitamins: Functions, Deficiencies and Food SourcesAssasination ClassroomNo ratings yet
- Lydia Hall's Care, Cure, Core Nursing TheoryDocument3 pagesLydia Hall's Care, Cure, Core Nursing TheoryAssasination ClassroomNo ratings yet
- Manual For Basic Procedure - UpdateDocument19 pagesManual For Basic Procedure - UpdateAssasination Classroom100% (1)
- Marcos - Er - Journal ReadingDocument1 pageMarcos - Er - Journal ReadingAssasination ClassroomNo ratings yet
- Marcos Er KetorolacDocument2 pagesMarcos Er KetorolacAssasination ClassroomNo ratings yet
- Unit 7.1 Nursing DiagnosingDocument2 pagesUnit 7.1 Nursing DiagnosingAssasination ClassroomNo ratings yet
- Marcos - Er - 13 AreasDocument2 pagesMarcos - Er - 13 AreasAssasination ClassroomNo ratings yet
- Marcos Er NCPDocument1 pageMarcos Er NCPAssasination ClassroomNo ratings yet
- Topic 1 - History of Mental HealthDocument23 pagesTopic 1 - History of Mental HealthAssasination ClassroomNo ratings yet
- Group Assignment May 11Document1 pageGroup Assignment May 11Assasination ClassroomNo ratings yet
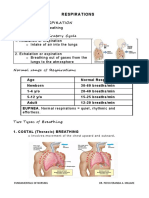
- Unit 5.2 RespirationsDocument5 pagesUnit 5.2 RespirationsAssasination ClassroomNo ratings yet
- Technique: Therapeutic Communication Examples RationaleDocument2 pagesTechnique: Therapeutic Communication Examples RationaleAssasination ClassroomNo ratings yet
- E Learning NCM 114Document2 pagesE Learning NCM 114Assasination ClassroomNo ratings yet
- 112 Midterm Elearning ANSwersDocument10 pages112 Midterm Elearning ANSwersAssasination ClassroomNo ratings yet
- Therapeutic: Communication Technique Examples RationaleDocument2 pagesTherapeutic: Communication Technique Examples RationaleAssasination ClassroomNo ratings yet
- Name:Synne Graile S. Marcos Grade and Section: 7 SincerityDocument1 pageName:Synne Graile S. Marcos Grade and Section: 7 SincerityArian May MarcosNo ratings yet
- BSN3C 2E Case Presentation On Blunt Abdominal Trauma November 2021Document42 pagesBSN3C 2E Case Presentation On Blunt Abdominal Trauma November 2021Assasination Classroom100% (1)
- Midterms CoparDocument4 pagesMidterms CoparAssasination ClassroomNo ratings yet
- Therapeutic communication techniquesDocument2 pagesTherapeutic communication techniquesAssasination ClassroomNo ratings yet
- Therapeutic Modalities 4th Edition Starkey Test BankDocument5 pagesTherapeutic Modalities 4th Edition Starkey Test BankGregoryGreenroyak100% (12)
- Covid19 Market Impact - OriginalDocument97 pagesCovid19 Market Impact - OriginalMUGARURA ALBERT BAKEBWANo ratings yet
- The Star News January 28, 2016Document36 pagesThe Star News January 28, 2016The Star NewsNo ratings yet
- JHA Conveyor Belt ChangingDocument9 pagesJHA Conveyor Belt ChangingMohammad Sharique PathanNo ratings yet
- Project Ashray - Planning A Time-Constrained ProjectDocument12 pagesProject Ashray - Planning A Time-Constrained ProjectShrey RanaNo ratings yet
- CNN International - Breaking News, US News, World News and VideoDocument8 pagesCNN International - Breaking News, US News, World News and VideomiculmucNo ratings yet
- Thyroid and Parathyroid Disorders: by Dr. Magdi Awad Sasi 2013Document65 pagesThyroid and Parathyroid Disorders: by Dr. Magdi Awad Sasi 2013Eidi IdhamNo ratings yet
- Sinew Surgical Technique PDFDocument8 pagesSinew Surgical Technique PDFLUNA SEQUOIANo ratings yet
- RPH Letter Biostatistics ReviewDocument14 pagesRPH Letter Biostatistics Reviewtpatel0986No ratings yet
- Vocabulary 2 KeperawatanDocument9 pagesVocabulary 2 KeperawatanSoull MechaniccNo ratings yet
- Eye InfectionsDocument13 pagesEye Infectionssara khanNo ratings yet
- Research - Understanding Genetics and Alzheimers DiseaseDocument6 pagesResearch - Understanding Genetics and Alzheimers DiseasetodoNo ratings yet
- DesmopressinDocument3 pagesDesmopressinKim RuizNo ratings yet
- Abilify Dosing GuideDocument2 pagesAbilify Dosing GuidemtassyNo ratings yet
- BiostatisticsDocument53 pagesBiostatisticshari rajanNo ratings yet
- Literature Review FinalDocument4 pagesLiterature Review Finalapi-549244735No ratings yet
- Activity: (If The Space Is Not Enough, You May Use The Blank Space at The Back of This Paper)Document2 pagesActivity: (If The Space Is Not Enough, You May Use The Blank Space at The Back of This Paper)Jordan CruzNo ratings yet
- TMJ DisordersDocument20 pagesTMJ DisordersDr. Hesham Zaatar100% (2)
- Tim Dosen: Nur Rachmat, Bpo Cica Trimandasari N, Amd Op Muhammad Syaifuddin, Amd OpDocument11 pagesTim Dosen: Nur Rachmat, Bpo Cica Trimandasari N, Amd Op Muhammad Syaifuddin, Amd OpAdi NugrohoNo ratings yet
- INBDEBooster Periodontics NotesDocument28 pagesINBDEBooster Periodontics NotesShawqi A100% (1)
- S 029 LBLDocument23 pagesS 029 LBLJabu MtoloNo ratings yet
- Platelets: Structure, Production, and DisordersDocument9 pagesPlatelets: Structure, Production, and DisordersShiena ArchividoNo ratings yet
- Health PlanningDocument46 pagesHealth PlanningJems Negn KeziewNo ratings yet
- Phlebotomy and Specimen ConsiderationsDocument21 pagesPhlebotomy and Specimen ConsiderationsLesly Marie LaxamanaNo ratings yet
- Health Education by Kamini MSNDocument15 pagesHealth Education by Kamini MSNkamini Choudhary100% (2)
- Common Cancers in WomenDocument49 pagesCommon Cancers in WomenNaghib BogereNo ratings yet
- Three Month Feeding Integration With Bifidobacterium Strains (Product Bifibaby) Prevents Gastrointestinal Symptoms in Healthy NewbornsDocument11 pagesThree Month Feeding Integration With Bifidobacterium Strains (Product Bifibaby) Prevents Gastrointestinal Symptoms in Healthy NewbornsLucas LE-DOANNo ratings yet
- Stroke (Bisaya and English)Document4 pagesStroke (Bisaya and English)AnaNo ratings yet